آخر المواضيع المضافة

 النبات
الحيوان
الأحياء المجهرية
علم الأمراض
التقانة الإحيائية
التقنية الحيوية المكروبية
التقنية الحياتية النانوية
علم الأجنة
الأحياء الجزيئي
علم وظائف الأعضاء
الغدد
المضادات الحيوية
النبات
الحيوان
الأحياء المجهرية
علم الأمراض
التقانة الإحيائية
التقنية الحيوية المكروبية
التقنية الحياتية النانوية
علم الأجنة
الأحياء الجزيئي
علم وظائف الأعضاء
الغدد
المضادات الحيوية| الوراثة الكمية Quantitative Genetics |


|
|
أقرأ أيضاً
التاريخ: 15-11-2015
التاريخ: 27-10-2015
التاريخ: 19-10-2020
التاريخ: 26-9-2018
|
الوراثة الكمية Quantitative Genetics
كانت الصفات المندلية الكلاسيكية صفات نوعية Qualitative trails سهلة التصنيف الى مجاميع متميزة من الأنماط الظاهرية ونتجت هذه الصفات من تأثير جين أو جينين بالأضافة الى الصفات النوعية يوجد الكثير من الصفات المهمة في النباتات والحيوانات والإنسان لا يمكن تصنيفها الى مجاميع متميزة . ولكن يمكن قياسها والتعبير عنها بوحدات قياس المسافة أو الوزن أو الحجم وبذا تشكل إختلافات مستمرة . وتكون هذه الصفات كمية Quantitative trails في طبيعتها وتنتج من فعل وتفاعل عدة جينات تتراوح من 10 – 100 جين أو أكثر من ذلك , وكذلك تتأثر هذه الصفات بعوامل البيئة المختلفة . ومن الأمثلة على الصفات الكمية هي الصفات المهمة زراعياً وأقتصادياً في الكثير من النباتات والحيوانات : كأنتاج الحبوب وإرتفاع النبات إنتاج الحليب والبيض . ولذا فإن معظم التجارب في المحطات الزراعية التجريبية في العراق وفي بقية أنحاء العالم تدور حول دراسة هذه الصفات وتحسينها . ومن الصفات الكمية في الأنسان هي القامة ووزن الجسم .
التوزيع الطبيعي للصفات الكمية Normal Distribution of Quantitative Trails
تظهر دراسة صفة كمية في عشيرة كبيرة بأن عدد قليل منالأفراد يحملون الأنماط الظاهرية القصوى , بينما أكثر الأفراد يكونون بالقرب من قيمة المعدل لتلك العشيرة . أن هذا النمط من التوزيع المتناظر يتصف بشكل الجرس ويطلق على هذا التوزيع بالتوزيع الطبيعي Normal distribution .
لاحظ علماء الوراثة بين عام 1900-1910م بإن الإختلافات المستمرة تعكس آلية وراثية تختلف عن تلك الأختلافات غير المستمرة , ووضعت فرضية الجينات المتعددة Multiple-gene Hypothesis لتفسير الإختلافات المستمرة . وجاء البرهان لهذه الفرضية من الأبحاث الكلاسيكية لنلسن أيلي Nilsson-Ehle في السويد وايست East في الولايات المتحدة للفترة بين 1910م الى 1913م .
لون بذرة الحنطة Seed Color of Wheat
وإحدى الدراسات التي أجراها نلسن تضمنت تضريبات بين صنف من الحنطة ذي بذور حمراء وآخر ذي بذور بيضاء . كانت بذور الجيل الأول ذات لون متوسط بين الأبوين , حيث كانت أفتح من البذور الحمراء للصنف الأبوي الأول ولكن أغمق من البذور البيضاء للصنف الأبوي الثاني . وعند ترتيب بذور الجيل الثاني حسب كثافة اللون , لوحظ تدرج مستمر من الأحمر الى الأبيض وكانت من بذور الجيل الثاني النسب : بحوالي 116 حمراء مثل الأب الأحمر البذور وبحوالي 116 بيضاء وبحوالي 1416 متوسط اللون أي تتراح بين لوني الأبوين . وعندما صنفت بذور الجيل الثاني ذات اللون المتوسط بصورة أدق على أساس كثافة اللون , شوهدت النسب : بحوالي 416ذات لون أعمق من لون بذور الجيل الأول وبحوالي 616 ذات لون متوسط مثل لون بذور الجيل الأول وبحوالي 416 ذات لون أفتح من لون بذور الجيل الأول . تدل هذه النتائج على إنعزال مستقل لزوجين من الجينات أو الجينات المضاعفة Duplicate genes التي تؤثر على نفس الصفة وذات تأثير متجمع . كذلك أجرى نلسن تضريبات أخرى بين صنفين آخرين من الحنطة : الأول ذو حبوب حمراء والثاني ذو حبوب بيضاء . كانت بذور الجيل الأول ذات لون متوسط .
ولكن بذور الجيل الثاني كانت نسبها حوالي 164 ذات بذور حمراء وحوالي 164 ذات بذور بيضاء , وحوالي 6264 ذات بذور متدرجة في اللون بين لوني الأبوين , وعندما صنفت هذه البذور بصورة أدق شوهدت النسب حوالي 664 ذات لون أحمر أفتح قليلاً من الأب الأحمر البذور , وحوالي 1564 ذات لون أفتح من لون بذور الجيل الأول , وحوالي 2064 ذات لون متوسط مثل لون بذور الجيل الأول وحوالي 1564 ذات لون أفتح من لون بذور الجيل الأول وحوالي 664 ذات لون أعمق قليلاً من بذور الأب الأبيض .
ونتائج هذا التضريب تشابه نتائج التضريب المندلي الثلاثي الهجين من حيث الأساس , ولذا فرض نلسن وجود ثلاثة أزواج من الجينات المستقلة لتفسير هذه النتائج بدلاً من زوجين في التضريب الأول , ومن الواضح بأن زوج واحد الذي أنعزل في التضريب الثاني كان متماثل الزيجة في كل من الأبوين في التضريب الأول .
طول كوز الذرة الصفراء Ear Length in Maize
درس ايست East عام 1913م وراثة طول كوز الذرة الصفراء بتضريب الصنف توم ثمب Tom Thumb قصير الكوز ذا مدى 5- 8 سم ومعدل 6.6سم بالصنف بلاك مكسيكان Black Mexican طويل الكوز ذا مدى 13-21 سم ومعدل 16.8 سم . كان طول كيزان الجيل الأول (F1) متوسط بين الأبوين وذا مدى 9-15 سم ومعدل 12.1سم .
أما طول كيزان الجيل الثاني (F2) فإنها بلغت مدى واسع (7-21سم) , وكان عدد قليل منها بقدر طول كيزان الصنف القصير الكوز, وعدد قليل آخر منها بقدر طول كيزان الصنف الطويل الكوز , وعدد كبير منها متوسطة الطول بقدر طول كيزان الجيل الأول بمعدل 12.9سم . إن هذه النتائج من حيث الأساس مشابهة الى نتائج لون بذر الحنطة , لكن في هذه الحالة لا يمكن تصنيف الكيزان الى مجاميع متميز من حيث الطول . الا إن الزيادة في الإختلاف في الجيل الثاني مقارنة بالجيل الأول يمكن تفسيرها على أساس انعزال عدد من الجينات التي تؤثر على طول الكوز بصورة تجميعية .وبذا يكون سبب الإختلاف بين كيزان كل من الأبوين والجيل الأول بيئي , أما سبب الإختلاف بين كيزان الجيل الثاني بيئياً ووراثياً (بسبب إنعزال عدد من الجينات التي قدرت بأربعة أزواج وكل زوج ينتج تأثير متساوي بمقدار 2.55 سم الى الطول الأساسي من الكوز) .
وتوجد أمثلة أخرى شائعة كلون البشرة في الإنسان وطول أوراق التويج في التبغ وحجم الأرنب التي توضح فرضية الجينات المتعددة . ويعتبر الآن مفهوم الجينات المتعددة للصفات الكمية أحد الأساسيات المهمة في علم الوراثة , حيث يدعم هذا المفهوم بإستعمال الطرق الإحصائية التي صممت من قبل فشر Fisher في إنكلترا ورايت Wright في الولايات المتحدة وغيرهم من علماء الوراثة . وترتكز وراثة الصفات الكمية على جينات كثيرة (متعددة) المنعزلة بإستقلال عادةً , ولكنها تؤثر على نفس النمط الظاهري وبطريقة تجميعية , وكل جين ينتج جزء من التأثير الكلي ولا توجد سيادة كاملة بين الأليلات وتؤثر عوامل البيئة على الناتج النهائي للصفات الكمية ويمكن التعبير عن النمط الظاهري للصفة الكمية بالمعادلة التالية :
النمط الظاهري = النمط الجيني + البيئة + (النمط الجيني × البيئة)
ويمكن قياس تأثير كل جزء من المعادلة إحصائياً بواسطة الإختلاف Variance وتصبح المعادلة :
الإختلاف بالنمط الظاهري= الإختلاف بالنمط الجيني + الإختلاف بالبيئة + الإختلاف (النمط الجيني × البيئة) , وبتعبير آخر :σ2p = σ2G + σ2E + σ2GE
وعند دراسة هذه الصفات يجب فصل التأثير الوراثي عن التأثير البيئي بإستعمال طرق أحصائية خاصة .
تقرير عدد الجينات للصفات الكمية Estimating the Number of Genes for Quantitative Traits
تساعد معرفة عدد الجينات التي تعين الصفات الكمية في تطوير طرق جديدة وكفوءة للبحث عنها , وكذلك يمكن الإستفادة من هذه المعلومات مباشرةً في تربية النبات والحيوان . ولكن يصعب تعيين عدد الجينات بالضبط المشمولة في تضريب ما بسبب وجود الإختلافات البيئية والإختلافات الوراثية بنفس القياس . ولبعض الصفات يمكن إجراء تقدير تقريبي بواسطة تعيين تردد حدوث كلاً من الطرفين في الجيل الثاني اللذين يشبهان النمط الظاهري للأبوين . وتعتبر هذه الطريقة طريقة تقريبية لعدد الجينات الفعالة ولكنها مفيدة للتحليل الوراثي الأولي .
وتوجد طريقة أخرى لتقدير عدد الجينات للصفة الكمية بالإستفادة من قيمة الإختلاف الوراثي Genotypic variance وتحت شروط معينة . تتألف عشيرة السلالة أو الخط النقي من أفراد متشابهة بالتركيب الوراثي الا انها تظهر الإختلاف بالنمط الظاهري , وعليه يكون كل هذا الإختلاف في الخط النقي إختلافاً بيئياً Environment variance وينتج من التضريبات بين صنفين نقيين هجين في الجيل الأول متناسق وراثياً وعليه يكون الإختلاف بالنمط الظاهري بين أفراد الجيل الأول إختلافاً بيئياً أيضاً , ويكون الإختلاف بين أفراد الجيل الثاني أكثر من الإختلاف بين أفراد الجيل الأول , بسبب الإنعزال وتكون تراكيب جديدة في أفراد الجيل الثاني . وفي الصفة الكمية ذات التوزيع الطبيعي تكون معدلات أفراد الجيل الأول والجيل الثاني متوسط بين معدلي الأبوين . وإذا لم يحدث تبدل في البيئة من جيل لآخر (ويمكن السيطرة على ذلك بالتجربة) فإن إختلاف البيئة بين أفراد الجيل الأول يجب أن يكون مساوياً الى إختلاف البيئة بين أفراد الجيل الثاني . لذا تعزى الزيادة في الإختلاف للنمط الظاهري بين أفراد الجيل الثاني على الإختلاف للنمط الظاهري بين أفراد الجيل الأول الى أسباب وراثية . وعليه فإن الإختلاف الوراثي بين أفراد الجيل الثاني (σ2GF2) يساوي الإختلاف للنط الظاهري بين أفراد الجيل الثاني (σ2PF2) ناقصاً الإختلاف للنمط الظاهري بين أفراد الجيل الأول (σ2PF1) , ويمكن التعبير عن ذلك بالمعادلة التالية :σ2GF2 = σ2PF2 - σ2PF1
ويعبر عن الإختلاف الوراثي بين أفراد الجيل الثاني بالمعادلة التالية :σ2GF2 = (a2N) / 2
حيث ترمز a الى مقدار مساهمة كل أليل فعال و N ترمز الى عدد أزواج الجينات المتعلقة في الصفة الكمية . ويمكن تقدير قيمة a من الصيغة : a = D/2N حيث ترمز D الى قيمة الفرق بين معدلي الأبوين لصفة كمية وبالتعويض يكون مقدار N كما يلي :σ2PF2 - σ2PF1 = σ2GF2 = a2N /2 = D2 / 8N
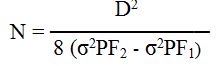
يرمز σ2PF1 الى الإختلاف بالنمط الظاهري بين أفراد الجيل الأول .
ويرمز σ2PF2 الى الإختلاف بالنمط الظاهري بين أفراد الجيل الثاني .
ويرمز σ2GF2 الى الإختلاف بالنمط الوراثي بين أفراد الجيل الثاني .
وعند إستعمال المعادلة الأخيرة يجب توفر الشروط (الفروض) التالية :
طبيعة الجينات التي تؤثر على الصفات الكمية The Nature of the Genes Affecting Quantitative Traits
سبق وإن قدمنا فرضية الجينات المتعددة Multiple-gene hypothesis لتفسير وراثة الصفات الكمية والتي تضمنت وجود عدة جينات واقعة على الكروموسومات . إن هذه الجينات مشابهة لبقية الجينات التي درسناها سابقاً في كل النواحي . ما عدا إن تأثيرها الفردي على النمط الظاهري قليل وتتأثر بالبيئة . أطلق ماذر Mather على هذه الجينات " بالجينات المتعددة " Polygenes ، وبذا فإنه ميزها عن الجينات الرئيسية Major genes التي يمكن تشخيصها بوضوح بسبب تأثيراتها الجلية المفردة . ويدل تقديم هذان المصطلحان على وجود التداخل بين المجموعتين الذي يمكن توضيحه بالنقاط التالية :
1- التأثيرات المضاعفة الناتجة من أنظمة الجينات المتعددة والرئيسية :
قد يتضاعف تأثير النمط الظاهري الناتج من الجينات المتعددة بفعل جين رئيسي واحد فمثلاً يرى المدى الطبيعي المستمر لقامة الإنسان بأنها تعتمد على تأثيرات الجينات المتعددة وقد تعزى القامة لشخص قصير جداً إلى تأثيرات الكثير من الجينات المختلفة ، ولكن وجود جين رئيسي واحد لقامة القزم قد تؤدي إلى نفس النتيجة .
2- الدور الممكن للجينات الرئيسية في أنظمة الجينات المتعددة :
يوجد أدلة على إن بعض الجينات تؤثر على كل من الصفات النوعية والكمية بنفس الوقت ، ففي البرسيم الأبيض مثلاً يتفاعل جينان سائدان ومستقلان لإنتاج التبقع Mottling في نصل الورقة الذي يعتبر صفة نوعية . ولكن جرعة هذين الجينين السائدين لها تأثير واضح على عدد الأوراق الذي يعتبر صفة كمية في البرسيم الأبيض .
3- الإرتباط بين الجينات المتعددة والجينات الرئيسية :
من الأمثلة على الإرتباط بين الجينات الرئيسية والجينات المتعددة هو إرتباط لون الفاصوليا (البذرة) المعين لجين واحد مع وزن الفاصوليا التي هي صفة كمية . وكذلك يرتبط لون ثمرة الطماطا بحجمها . وفي ذبابة الفاكهة تظهر جينات رئيسية في الكروموسومات الأربعة إرتباطاً مع جينات مختلفة لحجم البيضة التي تعتبر صفة كمية . وفي الفأر المنزلي يرتبط جين لون الفرو الأسمر مع عدد من الجينات التي تعين وزن البالغ وطول العظم في الذراع الخلفي .
التوريث Heritability
تعرف التوريث بأنها درجة سيطرة الوراثة على صفة معينة . ومعرفة مقدارها لكل صفة كمية ضروري لإتباع طريقة معينة في التربية وتحسين هذه الصفة . ويرمز لها بالرمز h2 والتي تساوي نسبة الإختلاف بالنمط الوراثي σ2G إلى الإختلاف بالنمط الظاهري σ2P أي إن :
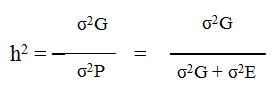
وقد يمكن قياس التوريث لكل صفة كمية على مستويين : الأول يكون واسعاً Broad Sense الذي يتضمن كل الإختلاف الوراثي الناتج من التأثير الإضافي (A) Additive للجينات المتعددة وتأثير السيادة (D) Dominance بين أليلات الجينات المتعددة وتأثير التفاعل بين الجينات المتعددة (I) Interaction . والثاني يكون دقيقاً Narrow sense الذي يشمل الإختلاف الوراثي الناتج من التأثير الإضافي Additive فقط للجينات المتعددة .
وتكون قيمة التوريث من صفر إلى واحد . ويمكن حساب قيمة التوريث بالحس الواقع Board sense في الأمثلة التالية :
ويكون σ2G = 0وعليه تكون قيمة التوريث مساوية إلى الصفر .



|
|
|
|
لخفض ضغط الدم.. دراسة تحدد "تمارين مهمة"
|
|
|
|
|
|
|
طال انتظارها.. ميزة جديدة من "واتساب" تعزز الخصوصية
|
|
|
|
|
|
|
مشاتل الكفيل تزيّن مجمّع أبي الفضل العبّاس (عليه السلام) بالورد استعدادًا لحفل التخرج المركزي
|
|
|