آخر المواضيع المضافة

 النبات
الحيوان
الأحياء المجهرية
علم الأمراض
التقانة الإحيائية
التقنية الحيوية المكروبية
التقنية الحياتية النانوية
علم الأجنة
الأحياء الجزيئي
علم وظائف الأعضاء
الغدد
المضادات الحيوية
النبات
الحيوان
الأحياء المجهرية
علم الأمراض
التقانة الإحيائية
التقنية الحيوية المكروبية
التقنية الحياتية النانوية
علم الأجنة
الأحياء الجزيئي
علم وظائف الأعضاء
الغدد
المضادات الحيوية| Ribosomes Have Several Active Centers |


|
|
Read More
Date: 1-4-2021
Date: 28-10-2020
Date: 1-1-2016
|
Ribosomes Have Several Active Centers
KEY CONCEPTS
- Interactions involving rRNA are a key part of ribosome function.
- The environment of the tRNA-binding sites is largely determined by rRNA.
The basic ribosomal feature is that it is a cooperative structure that depends on changes in the relationships among its active sites during translation. The active sites are not small, discrete regions like the active centers of enzymes. Rather, they are large regions whose construction and activities may depend just as much on the rRNA as on the ribosomal proteins. The crystal structures of the individual subunits and bacterial ribosomes give us a good impression of the overall organization and emphasize the role of the rRNA. The 2.8 Å–resolution structure clearly identifies the locations of the tRNAs and the functional sites. Many ribosomal functions can now be accounted for in terms of its structure.
Ribosomal functions are centered around the interactions with tRNAs. Figure 1 shows the 70S ribosome with the positions of tRNAs in the three binding sites. The tRNAs in the A and P sites
are nearly parallel to one another. All three tRNAs are aligned with their anticodon loops bound to the mRNA in the groove on the 30S subunit. The rest of each tRNA is bound to the 50S subunit. The environment surrounding each tRNA is mostly provided by rRNA. In each site, the rRNA contacts the tRNA at parts of the structure that are universally conserved.
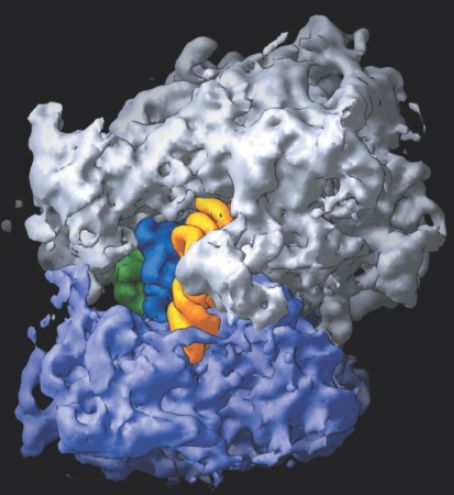
FIGURE 1. The 70S ribosome consists of the 50S subunit (white) and the 30S subunit (purple), with three tRNAs located superficially: yellow in the A site, blue in the P site, and green in the E site.
Photo courtesy of Harry Noller, University of California, Santa Cruz.
Before a high-resolution structure of the ribosome was available, it was a puzzle to understand how two bulky tRNAs could fit next to one another in reading adjacent codons. The crystal structure shows a 45° kink in the mRNA between the P and A sites, which allows the tRNAs to fit, as shown in the expansion of Figure 2.
The tRNAs in the P and A sites are angled at 26° relative to each other at their anticodons. The closest approach between the backbones of the tRNAs occurs at the 3′ ends, where they converge to within 5 Å (perpendicular to the plane of the page). This allows the peptide chain to be transferred from the peptidyltRNA in the P site to the aminoacyl-tRNA in the A site.
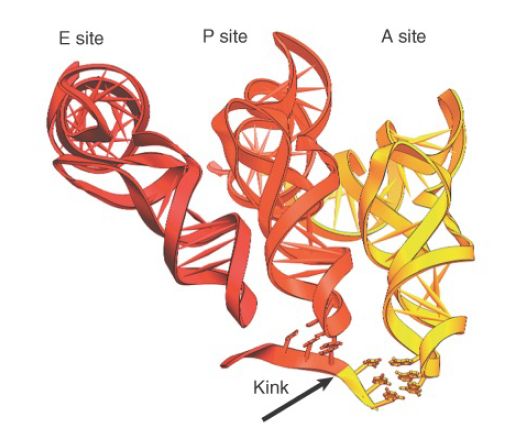
FIGURE 2. Three tRNAs have different orientations on the ribosome. mRNA turns between the P and A sites to allow aminoacyl-tRNAs to bind adjacent codons.
Photo courtesy of Harry Noller, University of California, Santa Cruz.
Aminoacyl-tRNA is inserted into the A site by EF-Tu, and its pairing with the codon is necessary for EF-Tu to hydrolyze GTP and be released from the ribosome (see the section earlier in this chapter titled Elongation Factor Tu Loads Aminoacyl-tRNA into the A Site). EF-Tu initially places the aminoacyl-tRNA into the small subunit, where the anticodon pairs with the codon. Movement of the tRNA is required to bring it fully into the A site, when its 3′ end enters the peptidyl transferase center on the large subunit. Different models have been proposed for how this process may occur. One suggests that the entire tRNA swivels so that the elbow in the Lshaped structure made by the D and TψC arms moves into the ribosome, enabling the TψC arm to pair with rRNA. Another suggests that the internal structure of the tRNA changes, using the anticodon loop as a hinge, with the rest of the tRNA rotating from a position in which it is stacked on the 3′ side of the anticodon loop to one in which it is stacked on the 5′ side. Following the transition, EF-Tu hydrolyzes GTP, allowing peptide bond formation to proceed.
Translocation involves large movements in the positions of the tRNAs within the ribosome. The anticodon end of tRNA moves about 28 Å from the A site to the P site, and then moves an
additional 20 Å from the P site to the E site. As a result of the angle of each tRNA relative to the anticodon, the bulk of the tRNA moves much larger distances: 40 Å from the A site to the P site and 55 Å from the P site to the E site. This suggests that translocation requires a major reorganization of structure. For many years, it was thought that translocation could occur only
in the presence of the factor EF-G. However, the antibiotic sparsomycin (which inhibits peptidyl transferase activity) triggers translocation. This suggests that the energy to drive translocation is actually stored in the ribosome after peptide bond formation has occurred. Usually EF-G acts on the ribosome to release this energy and enable it to drive translocation, but sparsomycin can play the same role. Sparsomycin inhibits peptidyl transferase by binding to the peptidyl-tRNA, blocking its interaction with aminoacyltRNA.
It probably creates a conformation that resembles the usual posttranslocation conformation, which, in turn, promotes movement of the peptidyl-tRNA. The conclusion is that translocation is an intrinsic property of the ribosome.
The hybrid state model suggests that translocation may take place in two stages, with one ribosomal subunit moving relative to the other to create an intermediate stage in which there are hybrid tRNA-binding sites (50S E/30S P and 50S P/30S A). Comparisons of the ribosome structure between pre- and posttranslocation states, and comparisons in 16S rRNA conformation between free 30S subunits and 70S ribosomes, suggest that mobility of structure is especially marked in the head and platform regions of the 30S subunit. An interesting insight into the hybrid state model is provided by the fact that many bases in rRNA involved in subunit
association are close to bases involved in interacting with tRNA.
This suggests that tRNA-binding sites are close to the interface between subunits and carries the implication that changes in subunit interaction could be connected with movement of tRNA. Much of the structure of the bacterial ribosome is occupied by its active centers. The schematic view of the ribosomal sites in Figure 3. shows they comprise about two-thirds of the ribosomal structure. A tRNA enters the A site, is transferred by translocation into the P site, and then leaves the ribosome by the E site. The A and P sites extend across both ribosome subunits; tRNA is paired with mRNA in the 30S subunit, but peptide transfer takes place in the 50S subunit. The A and P sites are adjacent, enabling translocation to move the tRNA from one site into the other. The E site is located near the P site (representing a position en route to the surface of the 50S subunit). The peptidyl transferase center is located on the 50S subunit, close to the aminoacyl ends of the tRNAs in the A and P sites (see the next section, 16S rRNA Plays
an Active Role in Translation).
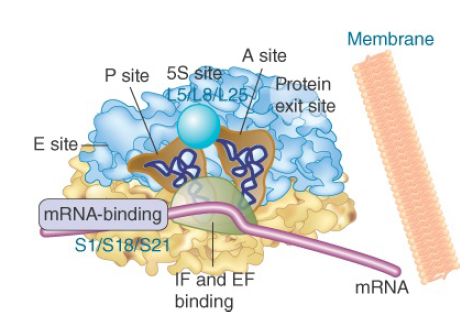
FIGURE 3. The ribosome has several active centers. It may be associated with a membrane. mRNA takes a turn as it passes through the A and P sites, which are angled with regard to each other. The E site lies beyond the P site. The peptidyl transferase site (not shown) stretches across the tops of the A and P sites. Part of the site bound by EF-Tu/G lies at the base of the A and P sites.
All of the GTP-binding proteins that function in translation (EF-Tu,EF-G, IF-2, RF1, RF2, and RF3) bind to the same factor-binding site (sometimes called the GTPase center), which probably triggers their hydrolysis of GTP. This site is located at the base of the stalk of the large subunit, which consists of the proteins L7 and L12. (L7 is a modification of L12 and has an acetyl group on the Nterminus.)
In addition to this region, the complex of protein L11 with a 58-base stretch of 23S rRNA provides the binding site for some antibiotics that affect GTPase activity. Neither of these ribosomal structures actually possesses GTPase activity, but they are both necessary for it. The role of the ribosome is to trigger GTP hydrolysis by factors bound in the factor-binding site.
Initial binding of 30S subunits to mRNA requires protein S1, which has a strong affinity for single-stranded nucleic acid. It is responsible for maintaining the single-stranded state in mRNA that is bound to the 30S subunit. This action is necessary to prevent the mRNA from taking up a base-paired conformation that would be unsuitable for translation. S1 has an extremely elongated structure and associates with S18 and S21. The three proteins constitute a domain that is involved in the initial binding of mRNA and in binding initiator tRNA. This locates the mRNA-binding site in the vicinity of the cleft of the small subunit. The 3′ end of rRNA, which pairs with the mRNA initiation site, is located in this region.
The initiation factors bind in the same region of the ribosome. IF-3 can be crosslinked to the 3′ end of the rRNA, as well as to several ribosomal proteins, including those probably involved in binding mRNA. The role of IF-3 could be to stabilize mRNA–30S subunit binding; then it would be displaced when the 50S subunit joins. The incorporation of 5S RNA into 50S subunits that are assembled in vitro depends on the ability of three proteins—L5, L8, and L25—to form a stoichiometric complex with it. The complex can bind to 23S rRNA, although none of the isolated components can do so. It lies in the vicinity of the P and A sites.
A nascent polypeptide extends through the ribosome, away from the active sites, into the region in which ribosomes may be attached to membranes. A polypeptide chain emerges from the ribosome through an exit channel, which leads from the peptidyl transferase site to the surface of the 50S subunit. The tunnel is composed mostly of rRNA. It is quite narrow—only 1 to 2 nm wide —and is about 10 nm long. The nascent polypeptide emerges from the ribosome about 15 Å away from the peptidyl transferase site. The tunnel can hold about 50 amino acids and probably constrains the polypeptide chain so that it cannot completely fold until it leaves the exit domain, though some limited secondary structures may form.



|
|
|
|
"عادة ليلية" قد تكون المفتاح للوقاية من الخرف
|
|
|
|
|
|
|
ممتص الصدمات: طريقة عمله وأهميته وأبرز علامات تلفه
|
|
|
|
|
|
|
الأمين العام للعتبة العسكرية المقدسة يستقبل شيوخ ووجهاء عشيرة البو بدري في مدينة سامراء
|
|
|