آخر المواضيع المضافة

 النبات
الحيوان
الأحياء المجهرية
علم الأمراض
التقانة الإحيائية
التقنية الحيوية المكروبية
التقنية الحياتية النانوية
علم الأجنة
الأحياء الجزيئي
علم وظائف الأعضاء
الغدد
المضادات الحيوية
النبات
الحيوان
الأحياء المجهرية
علم الأمراض
التقانة الإحيائية
التقنية الحيوية المكروبية
التقنية الحياتية النانوية
علم الأجنة
الأحياء الجزيئي
علم وظائف الأعضاء
الغدد
المضادات الحيوية| Internal Organization of Stems |


|
|
Read More
Date: 9-11-2016
Date: 27-10-2015
Date: 19-10-2016
|
Internal Organization of Stems
THE ARRANGEMENT OF PRIMARY TISSUES
In order to function properly, a tissue must contain the right cells in the proper arrangement. The same principle is true on a larger scale: In order to function properly, the tissues of an organ must be arranged correctly.
Epidermis. The outermost surface of an herbaceous stem is the epidermis, a layer of parenchyma cells (Fig. 1). Their outer tangential walls are the interface between the plant and the environment and regulate the interchange of material between the plant and its surroundings.
Because air is almost always drier than living cells, even when the relative humidity is high, preventing the loss of water to the air is critical for land plants. Also, the epidermis is a barrier against invasion by bacteria and fungi as well as small insects. The epidermis shields delicate internal cells from abrasion by dust particles, passing animals, or leaves and stems that might rub together, and its reflectivity protects the plant from overheating in bright sunlight.
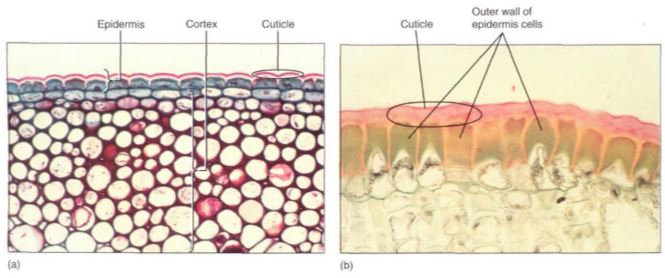
FIGURE 1 (a) The cuticle on this ivy (Hedera helix) has been stained dark pink; the lighter layer is wall material impregnated with cutin. The epidermal protoplasts have stained dark blue. Interior cells are photosynthetic collenchyma (X 120). b) This epidermis was taken from a desert plant, Agave. Its cuticle is much thicker than that of ivy in (a), and the outer walls of the epidermal cells have become extremely thick. The thickness of both the wall and cuticle strengthens the leaf surface against insects and prevents water loss (X 200).
The outer walls are encrusted with cutin, a fatty substance that makes the wall impermeable to water (Fig. 1). In species that occur in mild, moist habitats, such encrustation may be sufficient for water retention, but in most plants cutin builds up as a more or less pure layer called the cuticle. Under more severe conditions, even a cuticle is not sufficient, and a layer of wax may be present outside of the cuticle (Fig. 2; Table 1). Cutin and wax provide defense against pathogens like fungi and bacteria because digestive enzymes secreted by these microbes cannot break down either cutin or wax. In some species, the cuticle is so smooth that fungal spores cannot even stick to it and are washed off by rain or shaken off by wind. Waxes are indigestible and non-nutritious, so a thick layer of wax makes it difficult and unrewarding for an insect to chew into a stem.
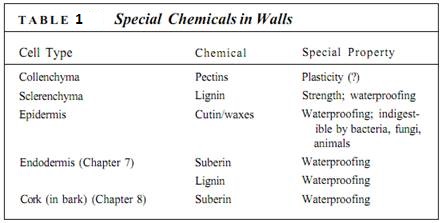

FIGURE 2: On this epidermis, wax is present as flat plates, but in other species it can occur as threads, beads, flakes, or a liquid (X 550).
Unfortunately, cutin and wax also inhibit the entry of carbon dioxide needed for photosynthesis—a totally impermeable epidermis would lead to the plant's starvation. However, the epidermis contains pairs of cells (guard cells) with a hole (stomatal pore) between them (Fig. 3). Guard cells and a stomatal pore together constitute a stoma (plural: stomata). Stomatal pores can be opened during the daytime, permitting carbon dioxide to enter the plant.
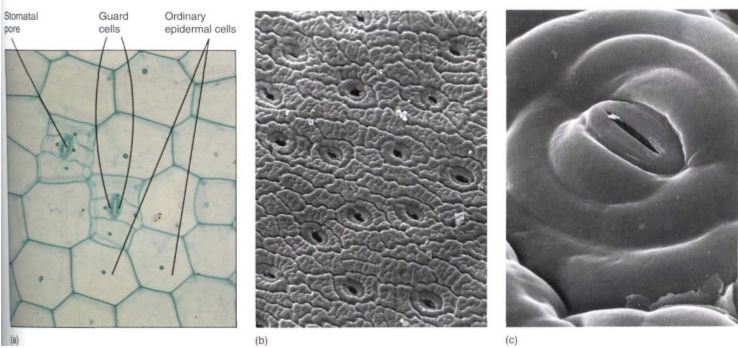
FIGURE 3: (a) Light microscope view of the epidermis of Zebrina; the stomatal pore, guard cells, and adjacent cells are visible, as are ordinary epidermal cells (X 150). (b) Scanning electron micrograph of the epidermis of a cactus showing numerous stomata. Notice how close together the stomata are; no chlorenchyma cell inside the cactus (on the other side of the epidermis) is far from a point where carbon dioxide can enter (X 50). (c) The stomatal pore is surrounded by guard cells, then other epidermal cells. Notice the small particle of debris at one end of the pore; clogging by debris is a problem in dusty areas (X 800).
The opening of the stomatal pore is made possible by the structure of the guard cells. The walls of the guard cells have a special radial arrangement of cellulose microfibrils which causes some parts to be weaker and more extensible than others. Guard cells swell by absorbing water; the walls next to the stomatal pore are rigid and do not extend, but the back walls are weak and stretch (Fig. 4), causing the cells to form an arc and push apart, opening the pore and allowing the entry of carbon dioxide and the exit of oxygen by diffusion. This unavoidably allows water vapor to escape. After sunset, when photosynthesis is impossible, it is not advantageous to keep the stomata open; guard cells in most species shrink and collapse toward each other, closing the pore and preventing water loss.
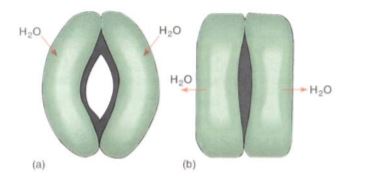
FIGURE 4: (a) When guard cells absorb water and swell, they become curved and the inner walls are pulled apart by the expansion of the back walls. (b) If guard cells lose water and shrink, they straighten, closing the stomatal pore.
In most plants, some epidermal cells elongate outward and become trichomes, also called hairs (Fig. 5). Trichomes make it difficult for an animal to land on, walk on, or chew into a leaf. They can shade the leaf by blocking some of the incoming sunlight, which may be too intense in summer. Trichomes can also create a layer of immobile air next to the leaf surface which allows water molecules that diffuse out of a stoma to bounce back in rather than be swept away by air currents.
Trichomes exist in hundreds of sizes and shapes; many are unicellular, being just long, narrow epidermal cells. Multicellular ones may be a single row of cells or several cells wide, and many branch in elaborate patterns. Most trichomes die shortly after maturity and their cell walls provide protection, but others remain alive and act as small secretory glands. Some secrete excess salt, others produce antiherbivore compounds, and those in the carnivorous plants secrete digestive enzymes onto the trapped insects. The poisonous, irritating compounds of stinging nettle are held in trichomes.
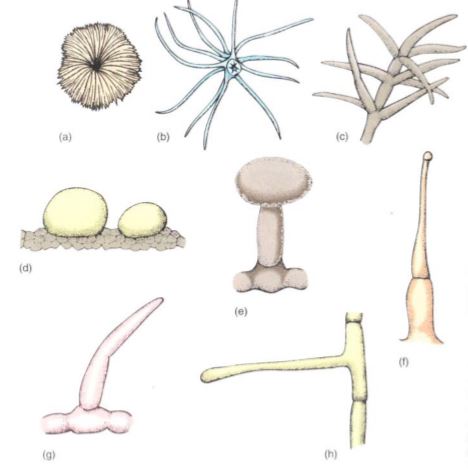
FIGURE 5: Trichomes. (a) Flat, scale-shaped trichome. (b) and (c) Branched trichomes. (d) Glandular trichomes—these are two large single cells. (e) Glandular trichome with a stalk cell elevating the secretory head cell. (f) Glandular trichome, capable of injecting poison. (g) and (h) Simple, unbranched, nonglandular trichomes.
Cortex. Interior to the epidermis is the cortex (Fig. 6). In many plants it is quite simple and homogeneous, composed of photosynthetic parenchyma and sometimes collenchyma. In other species it can be very complex, containing many specialized cells that secrete latex, mucilage, or pitch (resin). Some cortex cells contain large crystals of calcium oxalate or deposits of silica.
Cortex cells of most plants fit together compactly, but in fleshy stems such as tubers, corms, and succulents, the cortex parenchyma is aerenchyma, an open tissue with large intercellular air spaces. A few angiosperms, such as water lilies and the plants grown in aquaria, have become aquatic, living submerged in lakes or oceans. These plants have large cortical air chambers that provide buoyancy. The stems have such a strong tendency to float that no sclerenchyma is necessary for support.
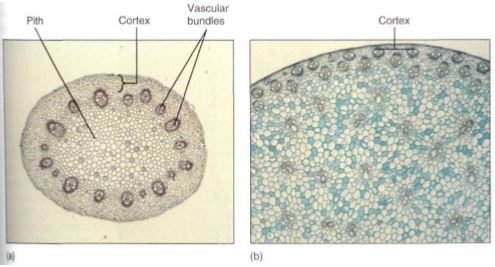
FIGURE 6: (a) The cortex of this stem of buttercup (Ranunculus) is the narrow band of cells between the epidermis and the vascular bundles (X 20). (b) The cortex of this corn stem is even narrower, in some places being only two or three cells wide (X 20).
Vascular Tissues. For very small organisms whose bodies are either unicellular or just thin sheets of cells, diffusion is adequate for the distribution of sugars, minerals, oxygen, and carbon dioxide throughout the body. But if any cells of the organism are separated from environmental nutrients and oxygen by just four or five cell layers, diffusion is too slow and a vascular system is necessary. Two types of vascular tissues occur in plants: xylem, which conducts water and minerals, and phloem, which distributes sugars and minerals.
The vascular systems of plants are quite different from those of animals. The plant vascular system is not a circulatory system: Water and minerals enter the xylem in the roots and are conducted upward to the leaves and stems. During its passage the xylem sap travels through dead cells, not through tubes composed of living cells like blood vessels. Once in the shoots, water evaporates from the surface of the leaves and is lost; the minerals are absorbed and used by surrounding cells. Phloem cells are living; they pick up sugar from areas where it is abundant, usually leaves during summer and tubers or rhizomes in spring, and transport it to areas where it is needed, especially the growing tips of shoots, roots, young leaves, and flowers. In the later months of summer, phloem carries sugar into developing fruits and into the storage organs of perennial plants. Because sugar must be dissolved to be conducted, water is transported simultaneously in phloem.
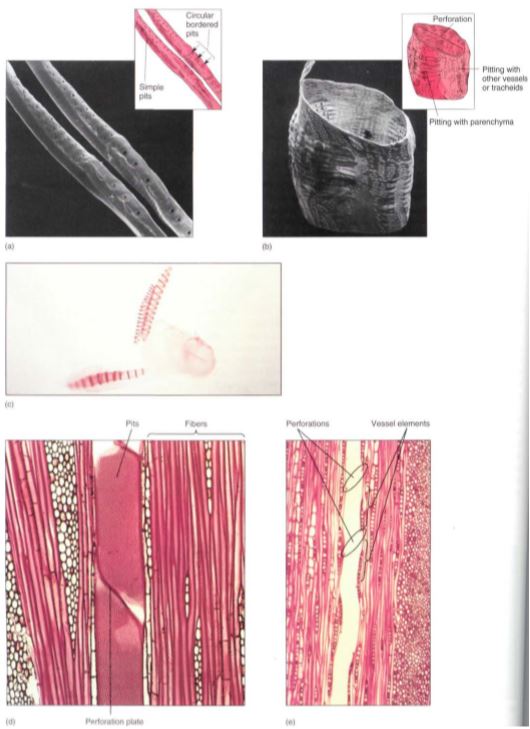
FIGURE 7: Tracheary elements. (a) and (c) Tracheids are long cells with tapered ends. The primary wall is complete over the whole surface, and in (c) it is so thin that the inner, spiral secondary wall is visible. In (a), the primary wall tore during specimen preparation, permitting views of circular bordered pits and simple pits. (b) and (d) Vessel elements tend to be wider and shorter than tracheids, but the most important feature is the perforation, the large hole at each end. (e) The perforation of one vessel element must be aligned with that of the next vessel element if water is to pass through with little friction; the stack of vessel elements is a vessel. The vessel here has been cut down the center, so the front wall and back wall are missing and the vessel appears to be just an open space, (a) X 600; (b) X 270; (c) X 100; (d) X 200; (e) X 50.
Xylem. Within xylem are two types of conducting cells: tracheids and vessel elements; both are types of sclerenchyma (Fig. 7; Table 2). The term "tracheary element" refers to either type of cell. As a young cell matures into a tracheary element, it first must enter cell cycle arrest and stop dividing. It is initially a small parenchyma cell with only a thin primary wall, but the cell becomes long and narrow and then deposits a secondary wall that reinforces the primary wall. The cell then dies and its protoplasm degenerates, leaving a hollow tubular wall (Fig. 8).
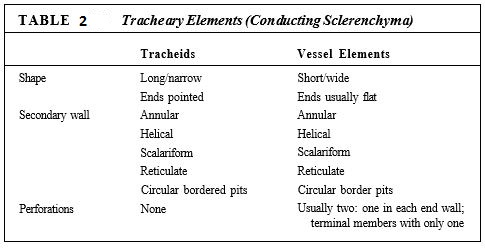
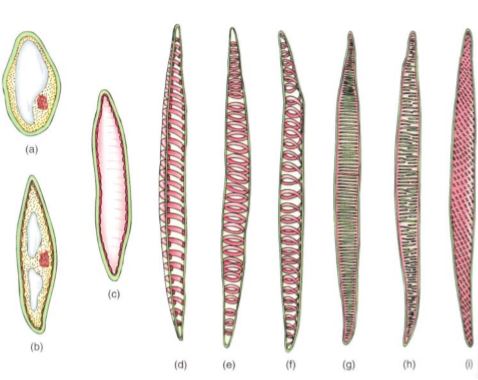
FIGURE 8: Tracheids. (a) to (d) The growth of a young cytoplasmic cell into a tracheid. Once the cell has almost reached mature size, the secondary wall is deposited interior to the primary wall (c). After the secondary wall is finished, the protoplasm dies and degenerates, leaving only the primary and secondary walls. (d) A tracheid cut open, showing a secondary wall in the form of helical thickenings. (e) A whole cell in which the primary wall is so thin that the annular secondary walls inside are visible. (f) Whole cell with helical secondary wall. (g) Scalariform secondary wall. (h) Reticulate secondary wall. (i) Pitted secondary wall.
The secondary wall is impermeable to water, so areas of the permeable primary wall must remain uncovered if water is to enter and leave the cell. In the simplest type of tracheary element, there is only a small amount of secondary wall, organized as a set of rings, called annular thickenings, on the interior face of the primary wall (Fig. 8e). This arrangement provides a large surface area for water movement into and out of the cell, but it does not provide much strength. The walls must be strong because the movement of the water tends to cause them to collapse inward (Fig. 9). With helical thickening, the secondary wall exists as one or two helices interior to the primary wall (see Fig. 8f). Scalariform thickening provides much more strength because the secondary wall covers most of the inner surface of the primary wall and is fairly extensive. Just as in non-conducting sclerenchyma, the area where secondary wall is absent is called a pit (Fig. 10a). In tracheary elements with reticulate thickening, the secondary wall is deposited in the shape of a net, as the name suggests. The most derived and strongest tracheary elements are those with circular bordered pits (Figs. 10b and 11). In such tracheary elements, virtually all the primary wall is. covered by the secondary wall. The pits that allow water movement are weak points in the wall, but the weakness is reduced by a border of extra wall material around the pit—hence the name.
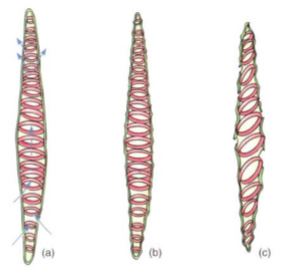
FIGURE 9: (a) As water enters a tracheary element, it passes through the primary wall between regions of secondary wall. (b) Water adheres to cellulose molecules, pulling the walls inward and upward as it rises in xylem. The primary wall tends to collapse under the friction of the passage, but the secondary wall provides strength to hold the primary wall in place. (c) Under severe water stress (dry air and dry soil), annular and helical walls may not be strong enough to prevent collapse.
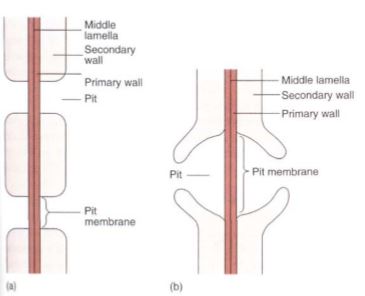
FIGURE10: (a) Side view of a pit that has been bisected. In a sclereid or fiber, the secondary wall would be thicker and the pit narrower and longer. The diameter of the pit can vary, being broad as in Figure 5. 10d or narrow as in Figure 5.12. (b) A bordered pit has a rim of extra wall material around it, which strengthens it and prevents the pit from being a weak spot in the cell. With borders, pits can be broad enough to allow rapid flow of water from tracheary element to tracheary element, but not so weak as to be detrimental. Only tracheary elements have bordered pits; fibers and sclereids do not.
Tracheary elements with annular thickenings are weak, but a large percentage of their primary wall is uncovered and available for water movement. Pitted tracheary elements are just the opposite: They are extremely strong, but so much of the surface is covered by secondary wall that water enters or leaves the cells slowly. Each is selectively advantageous under certain conditions and disadvantageous under others. If a species grows in perpetually wet soil and water moves easily, the cells are not in danger of collapsing, so tracheary elements with annular or helical secondary walls are adaptive, not only because the large open regions of primary wall allow water to pass through quickly but also because the plant minimizes the amount of sugar it spends building secondary walls. But if the soil dries even slightly, water is pulled with more force and exerts inward traction on the wall of the tracheary elements. The stronger scalariform, reticulate, or circular bordered pitted walls are necessary to keep the tracheary elements open.
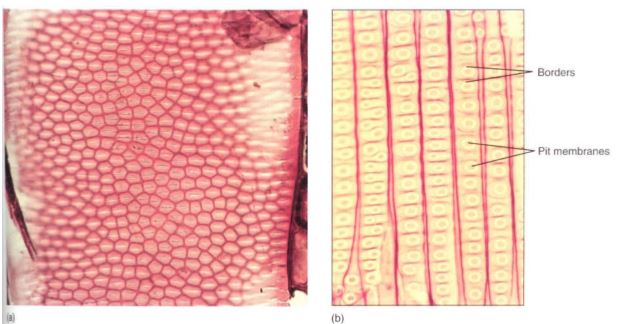
FIGURE 11: (a) Bordered pits in face view on front wall of a vessel element. The white slits are the actual openings of the pits, the pinkish area around each white region is the border, and the dark red hexagons are the regions where the secondary walls lie against the primary walls (X 400). (b) Bordered pits in face view on the front walls of tracheids of pine. This is the same magnification as (a); notice that tracheids are much narrower than vessel elements and that pits in pine are extremely large.
Although they cannot conduct water as quickly, water movement is automatically slower because the soil is dry. Under dry conditions, using extra glucose to build stronger secondary walls can be a valuable investment.
Tracheids obtain water from other tracheids below them and pass it on to those above; they must occur in groups, lying side by side with some and having their ends overlapping the ends of others (Fig. 12). Also, the pits of adjacent tracheids are aligned so that water can pass through. The aligned set of pits is a pit-pair, and the set of primary walls and middle lamella between them constitutes a pit membrane, just as in fibers and sclereids . Although very permeable to water, pit membranes do offer slight resistance. If each tracheid is 1 mm long, each water molecule passing from root to leaf in even a short plant of only 1 meter (1000 mm) must pass through 1000 pit membranes. The friction adds up. Vessel elements provide a way to move water with less friction. Vessel elements, like tracheids, are individual cells that produce both primary and secondary walls before they die at maturity. In vessel elements, however, an entire region of both primary and secondary wall is missing. During the final stages of differentiation, a large hole called a perforation is digested through a particular site of the primary wall, often removing the entire end wall . Because perforations are wide and have no pit membrane, they provide a pathway with little friction.
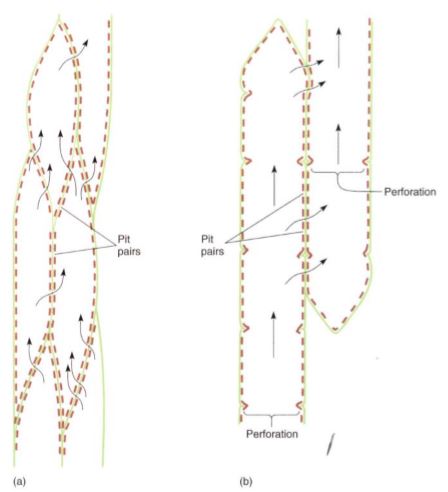
FIGURE 12: (a) Water can pass between tracheids only through pit-pairs. Other than plasmodesmata (which are too tiny to be important in water conduction), no holes are present in the primary walls of tracheids. (b) Water can pass between vessel elements through perforations, but it can pass from one vessel to another only through pit-pairs. Vessels are long, but not as long as a whole plant, so water must pass from one vessel to another as it moves upward from roots to stems to leaves. Green = primary wall; brown = secondary wall.
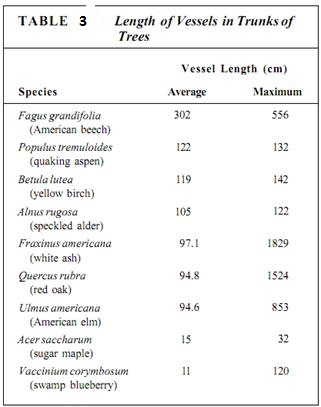
The perforations of adjacent vessel elements must be aligned, and each element must have at least two perforations, one on each end. An entire stack of vessel elements is a vessel. The vessel elements on each end of a vessel have only one perforation. Whereas vessel elements, being individual cells, are only about 50 to 100 µm long, vessels can be many meters long, running all the way from a root tip to a shoot tip, although some are only a few centimeters long (Table 3).
Vessels must absorb water from parenchymal cells, tracheids, or other vessels, and they must pass it on. Their side walls must have pits for this lateral transfer, and all the types of wall thickening mentioned above for tracheids also occur in vessel elements. The only constant difference is that vessel elements have perforations—complete holes—in their walls, whereas tracheids only have pits (see Table 2). The perforation greatly reduces the friction, and water moves much more easily through a set of vessels than through a set of tracheids.
All plants with vascular tissue have tracheids, which evolved at least 420 million years ago Vessels evolved more recently and occur almost exclusively in flowering plants, where they perform virtually all long-distance water conduction in roots and stems. In flowering plants, tracheids occur mostly in the fine veins of leaves but can also be found in the wood of some species. Almost all no-nangiosperms such as conifers and ferns have xylem com-posed exclusively of tracheids.
Phloem. Like xylem, phloem has two types of conducting cells, sieve cells and sieve tube members; the term "sieve element" refers to either one (Fig. 13). These are very different from tracheary elements: They must remain alive in order to conduct, and because they have only primary walls they are parenchyma cells. As immature sieve elements begin to differentiate, their plasmodesmata enlarge to a diameter of more than 1 /u.m (1000 nm) and are called sieve pores. Plasmodesmata occur in groups (primary pit fields), so sieve pores also occur clustered together in groups called sieve areas. Sieve elements remain alive during differentiation, and the plasma membrane that lined the plasmodesma continues to line the sieve pore (Fig. 14). The amount of cytoplasm within the pore also increases, and rapid, bulk movement from cell to cell becomes possible. Sieve pores of adjacent cells must be aligned.
The two types of sieve elements differ in shape and placement of sieve areas (Fig. 13). If the cell is elongate and spindle-shaped (like a tracheid) and has sieve areas distributed over all its surface, it is a sieve cell (Table 4). This type evolved first and is found in older fossils and in nonangiosperm vascular plants. Because phloem conducts mostly longitudinally, it is most efficient for the sieve areas on the two ends of the cell to be especially large and to have very wide sieve pores, whereas sieve areas on the sides of the cell can be rather small. Cells like this are sieve tube members; they are stacked end to end with their large sieve areas aligned, forming a sieve tube. These end-wall sieve areas with large sieve pores are sieve plates. Like vessels, sieve tubes are shorter than the plant, and the small, lateral sieve areas are important for transport between sieve tubes. Sieve tube members must have evolved at about the same time as the flower; all angiosperms have sieve tube members, but none of the nonangiosperms has them.
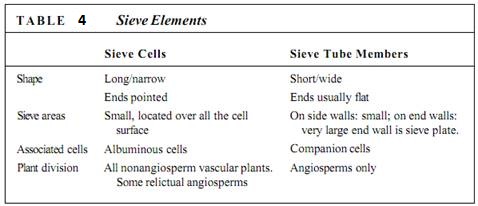
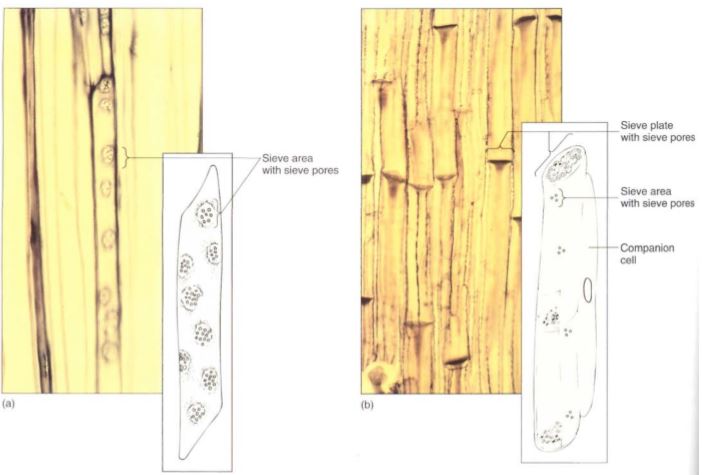
FIGURE 13: (a) A sieve cell is shaped like a tracheid and has sieve areas with sieve pores over much of its surface (X 250). (b) A sieve tube member has flat end walls (sieve plates) with large sieve pores. On the side walls are only a few small sieve areas with very narrow sieve pores (X 60). Note that the micrograph of sieve cells is a high magnification; they are much narrower than the sieve tubes of (b).
An unusual feature of sieve elements is that their nuclei degenerate but the cells remain alive. Cytoplasm cannot carry on complex metabolism without a nucleus, and in phloem the necessary nuclear control is provided by cells that are intimately associated with Ute conducting cells. Sieve cells are associated with albuminous cells, and sieve tube members are controlled by companion cells (Fig. 15).
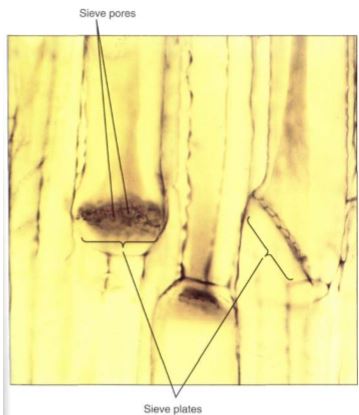
FIGURE 14: Large sieve areas present on the sieve plates of sieve tube members. Phloem sap flows from one tubelike sieve tube member into the next through the wide sieve pores (X 300).
These cells are often smaller than the accompanying conducting cell and have a prominent nucleus and a cytoplasm that is dense and filled with ribosomes. Walls between conducting cells and controlling cells have many complex passages that are sieve areas on the conducting cell side and large plasmodesmata on the controlling cell side.
Recent evidence shows that the companion cells have another important role, that of loading sugars into and out of the sieve tube members. The extent and importance of this [unction are unknown.
Vascular Bundles. Xylem and phloem occur together as vascular bundles, located just interior to the cortex (Fig. 16). In dicots, vascular bundles are arranged in one ring surrounding the pith, a region of parenchyma similar to the cortex. In monocots, vascular bundles are distributed as a complex network throughout the inner part of the stem; between the bundles are parenchyma cells. Monocot bundles are frequently described as "scattered" in the stem, suggesting that they occur at random, but they actually have precise, specific patterns that are too complex to be recognized easily.
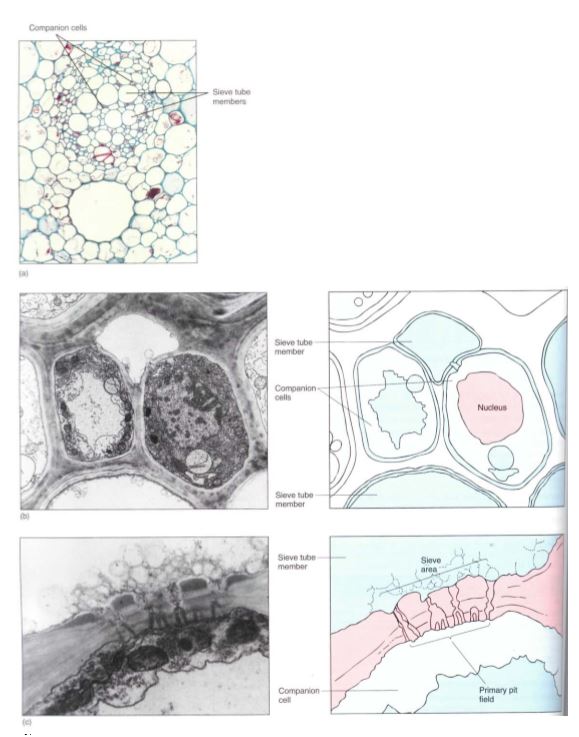
FIGURE 15: (a) In a vascular bundle, sieve tube members and companion cells occur together. Companion cells are small and rich in cytoplasm; sieve tube members are large and appear empty because most phloem sap is lost while preparing the tissue for microscopy (X 150). (b) Electron micrograph showing two companion cells and two sieve tube members. (c) The connection between a companion cell and a sieve tube member is a primary pit field with plasmodesmata on the side of the companion cell (bottom), but it is a sieve area with sieve pores on the side of the sieve tube members (top) (X 45,000).
All vascular bundles are collateral; that is, each contains both xylem and phloem strands running parallel to each other (Fig. 16). The xylem of a vascular bundle is primary xylem because it is part of the primary plant body. In addition to conductive tracheids and vessel elements, there is usually a large proportion of xylem parenchyma and even mechanical sclerenchyma in the form of xylem fibers. The vascular bundle phloem is primary phloem, and mixed with the sieve elements and companion cells or albuminous cells may be storage parenchyma and mechanical sclerenchyma, usually as phloem fibers, although phloem sclereids also occur. Cells of primary xylem are larger than cells of primary phloem, and the tracheary elements on the inner side of each bundle are much smaller than the outer ones (Fig. 16). This is true of the primary xylem in all stems, and; the reason for this becomes clear when the shoot growth is examined.
FIGURE 16: (a) In this vascular bundle of sunflower (Helianthus), the xylem contains rows of vessels alternating with rows of xylem parenchyma. The phloem contains sieve tube members and companion cells. There is a large sclerenchyma cap of phloem fibers (X 60). (b) The same bundle, shown at higher magnification (X 250). (c) The vascular bundles of monocots have a more complex arrangement. In this corn stem, many large bundles are located near the cortex and fewer, smaller bundles in the ground tissue (X 50). (d) High magnification of a corn bundle, showing primary xylem and phloem (X 250).



|
|
|
|
دراسة يابانية لتقليل مخاطر أمراض المواليد منخفضي الوزن
|
|
|
|
|
|
|
اكتشاف أكبر مرجان في العالم قبالة سواحل جزر سليمان
|
|
|
|
|
|
|
اتحاد كليات الطب الملكية البريطانية يشيد بالمستوى العلمي لطلبة جامعة العميد وبيئتها التعليمية
|
|
|