آخر المواضيع المضافة

 النبات
الحيوان
الأحياء المجهرية
علم الأمراض
التقانة الإحيائية
التقنية الحيوية المكروبية
التقنية الحياتية النانوية
علم الأجنة
الأحياء الجزيئي
علم وظائف الأعضاء
الغدد
المضادات الحيوية
النبات
الحيوان
الأحياء المجهرية
علم الأمراض
التقانة الإحيائية
التقنية الحيوية المكروبية
التقنية الحياتية النانوية
علم الأجنة
الأحياء الجزيئي
علم وظائف الأعضاء
الغدد
المضادات الحيوية| Secondary Xylem |


|
|
Read More
Date: 16-10-2016
Date: 30-10-2015
Date: 12-11-2016
|
Secondary Xylem
TYPES OF WOOD CELLS
All cells formed to the interior of the vascular cambium develop into secondary xylem, known as wood. The secondary xylem contains all the types of cells that occur in primary xylem but no new ones. Wood may contain tracheids, vessel elements, fibers, sclereids, and parenchyma. The only real differences between primary and secondary xylem are the origin and arrangement of cells. The arrangement of secondary xylem cells reflects that of the fusiform and ray initials: An axial system is derived from the fusiform initials and a radial system develops from the ray initials (Fig. 1).

FIGURE 1:Scanning electron micrograph of a cube of wood from a tulip tree (Liriodendron tulipifera). Examine the relationship between the various components, especially the large vessels and the ray parenchyma (X 110). (Courtesy of Dr. Wilfred A. Cote, Jr., State University of New York
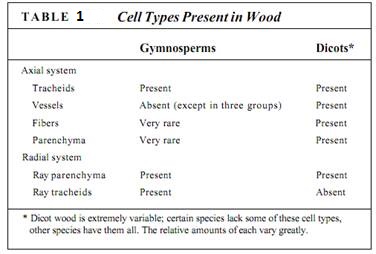
The axial system always contains tracheary elements (tracheids or vessels or both), which carry out longitudinal conduction of water through the wood (Fig. 2). In many species of dicots, the axial system also contains fibers that give the wood strength and flexibility (Fig. 3; Table 1). Most commercially important dicot woods contain large amounts of fibers, making them strong, tough, and useful for construction.
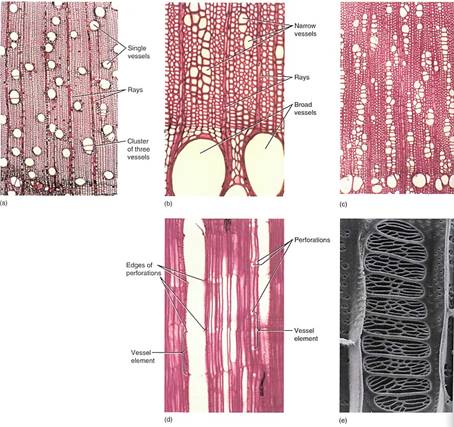
FIGURE 2:Vessels in wood. (a) In this wood, rather medium-size vessels occur in a mass of fibers. Each vessel occurs by itself or in a small cluster of two or three vessels (transverse section; X 40). (b) This wood has vessels of two very different sizes: extremely broad (at bottom of the micrograph) and very narrow (near the top). When water is plentiful, it can be transported in large quantities through the broad vessels. The narrow vessels are more effective at carrying water when the soil is dry and only a small amount of water is available to be transported (Cotinus americana, transverse section; X 150). (c) The vessels of the Pistasia mexicana are arranged in long radial groups. Because most vessels touch two or three other vessels, they can share water through the pits on their side walls (transverse section; X 50). (d) This longitudinal section passes through the lumen of one of the vessels. The front and back walls of the vessel elements were cut away during specimen preparation, so the vessel appears empty. The perforations are visible as short projections into the lumen (radial section; X 150). (e) Perforation between two vessel elements: The nearer cell has a scalariform perforation, and the farther cell has a reticulate perforation (X 700). (Courtesy of J. Ohtani, Hokkaido University)
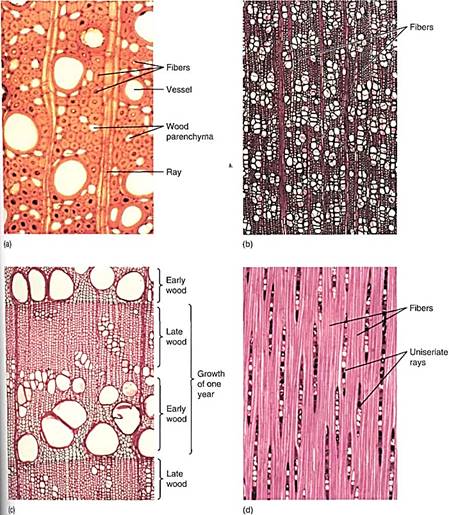
FIGURE 3:Wood fibers. (a) The fibers of this wood have extremely thick walls. Also, the fibers are abundant, so this wood is quite strong (transverse section; X 150). (b) Vessels make up over half the volume of this wood, so fibers are not very abundant. Furthermore, the fiber cell walls are rather thin; this wold not be a strong wood (transverse section; X 40). (c) In this wood, regions that have many large vessels and few fibers (called early wood, discussed later in this chapter) alternate with regions that have abundant fibers and only a few narrow vessels (late wood). Early wood provides maximum conduction capacity, whereas late wood provides strength (transverse section; X 50). (d) A longitudinal section through fiber-rich late wood. All fiber cells are long and narrow and have thick walls (tangential section; X 50).
They are called hardwoods, a term now used for all dicot woods, even those that lack fibers or are very soft, such as balsa. Woods from conifers such as pines and redwoods have few or no fibers and thus have a softer consistency. These are known as the softwoods, even though in many instances they are actually much harder than many hardwoods. Tracheary elements and fibers are elongate, as are the fusiform initials, but in many species, some immature cells undergo transverse divisions and differentiate into columns of xylem parenchyma (Fig. 4). Axial xylem parenchyma is important as a temporary reservoir of water; on cloudy or humid days and at night when leaves are losing little water, wood has a temporary surplus of conducting capacity and water is moved from roots into wood parenchyma and held there. When the air is hot and dry, leaves may lose water very rapidly; water can then be drawn from wood parenchyma if the soil is too dry to supply enough water. Conifers such as pine, cedar, juniper, and redwood have little or no axial parenchyma, so they have little reserve water. For them, tough, waxy, water-conserving leaves are selectively advantageous.
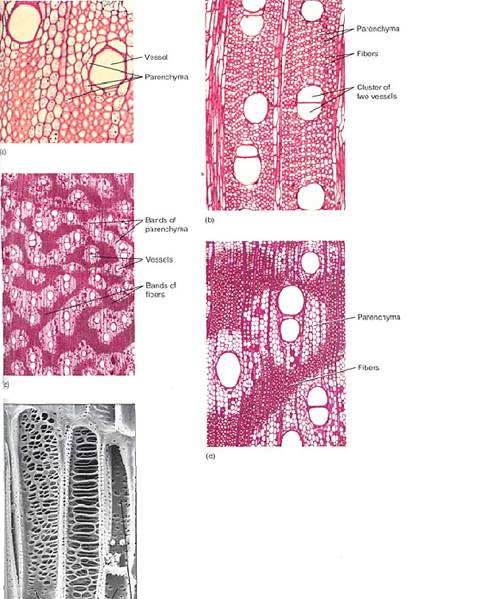
FIGURE 4:Wood axial parenchyma. (a) To prepare wood for microscopy, it is often boiled to make it soft enough to cut into thin specimens; unfortunately this destroys the protoplasm of any parenchyma cells. This wood was not boiled, so the starch-filled parenchyma cells are easily visible. The large cells surrounding the vessels are also parenchyma, and they can release water to the vessels in times of water shortage (temporarily dry soil) (transverse section; X 150). (b) This wood has abundant axial parenchyma—the small cells with thin walls. The small cells with thick walls are fiber cells. Because this wood was boiled, it is difficult to be certain which is parenchyma (transverse section; X 50). (c and d) The axial parenchyma in this wood forms large bands; notice that all the vessels occur in the bands, surrounded by parenchyma. None is in the fiber masses. This parenchyma may act as a "water jacket" around vessels, absorbing excess water when water loss from leaves is low (cool nights) and releasing it when water loss is rapid (hot days) (transverse sections; c, X 15; d, X 50). (e) Axial parenchyma cells adjacent to vessels (X 700). (Courtesy of J. Ohtani, Hokkaido University).
The complexity of the axial system of wood varies greatly. Most gymnosperms contain only tracheids in their axial systems; fibers and parenchyma cells are sparse or absent . Some dicots, especially the very relictual ones, have mostly just tracheids, but in most dicots all possible types of cells are present, and numerous cell-cell relationships are possible. Water-storing parenchyma may be immediately adjacent to vessels, or it can be arranged such that it never touches vessels. Fibers provide maximum strength if grouped together in masses, which is how they are usually arranged. If fibers are located around a vessel, their secondary walls reinforce the walls of the vessel and help it resist collapse. But the presence of fibers excludes parenchyma cells.
The radial system of xylem is much simpler. In flowering plants, it contains only parenchyma, arranged as uni-, bi-, or multiseriate masses called rays (see Figs. 1 and 5). Ray parenchyma cells store carbohydrates and other nutrients during dormant periods and conduct material over short distances radially within the wood. The two basic types of ray parenchyma cells are upright cells and procumbent cells (Fig. 5). At least in some plants, procumbent ray cells have no direct connection with axial cells, but upright cells do. The ray/axial interface can take many forms. If the upright ray parenchyma cell is adjacent to axial parenchyma, plasmodesmata occur. If the ray parenchyma is adjacent to an axial tracheid or vessel element, the tracheary element has pits in its secondary wall and the ray cell has very thin walls facing the pits. In early springtime, when trees such as maples are drawing on their nutrient reserves, the starch that had been stored in the upright cells is the first to be digested into sugar and passed into the axial tracheary elements for conduction to newly expanding buds, leaves, and flowers. Starch in procumbent cells is not digested until later and presumably must first be routed through upright cells for transfer to axial conducting cells.

FIGURE 5:Wood rays. (a) The rays in this wood are extremely broad, so they are easy to see. Notice that each ray extends from the xylem into the phloem (X 15). (b) These rays are uniseriate, only one cell wide; they are rather short, only about ten cells tall. This is from late wood, so vessels are not present in this section (tangential section; X 50). (c) These rays are multiseriate, several cells wide Over half the volume of this area of the wood is storage parenchyma, not conductive tracheary elements or strengthening fibers (tangential section; X 40). (d) This wood has giant multiseriate rays and small uniseriate rays (tangential section; X 50). (e) This ray has procumbent cells in the central part and upright cells along the edges (radial section; X 50). (f) Ray cells often contain crystals of stored material; four crystals are visible here in the cells along the margin. None occurs in the procumbent cells (radial section; X 150).
In gymnosperms, xylem rays are almost exclusively uniseriate; they are multiseriate only if they contain a resin canal (Fig. 6a). In addition to ray parenchyma cells, they may contain ray tracheids—horizontal, rectangular cells that look somewhat like parenchyma cells but have secondary walls, circular bordered pits, and protoplasts that degenerate quickly after the secondary wall is completed (Fig. 6b).

FIGURE 6: (a) Conifer rays often contain secretory canals that produce resin (pitch), important in preventing insects from burrowing through the wood (tangential section; X 150). (b) The central ray cells are living ray parenchyma, with large nuclei that have been stained black. The procumbent cells along the top and bottom of the ray are ray tracheids that have circular bordered pits but no protoplasm when mature (radial section; X 150).
ANNUAL RINGS
In regions with strongly seasonal climates, the vascular cambium is quiescent during times of stress, either winter cold or summer drought. But when quiescence ceases, the vascular cambium becomes active and cell division begins. At the same time, the new, expanding leaves are thin and delicate, and their cuticle is neither thick nor fully polymerized. Leaves like this lose water at a rapid rate, so trees need a high capacity for conduction at this time. The first wood formed is early wood, also called spring wood, and it must have a high proportion of wide vessels (Fig. 7a) or, in gymnosperms, wide tracheids (Fig. 7d). Later, the cuticle has thickened, transpiration is less, and large numbers of newly formed vessels are conducting rapidly. Wood produced at this time, called late wood or summer wood, can have a lower proportion of vessels. But the plant is a year older, it is larger and heavier, and it needs more mechanical strength to hold up the increased number of leaves and the larger branches. Late wood is stronger if it contains numerous fibers or, in gymnosperms, if it contains narrow, thick-walled tracheids. Finally, at the end of the growing season, the cambium becomes dormant again. The last cells often develop only as heavy fibers with especially thick secondary walls. In a tree with wood like that just described, it is easy to see late wood and early wood, the two together making up 1 year's growth, an annual ring.
An alternative arrangement exists for wood cells: In some species, vessels form throughout the growing season. Those produced in spring are neither more abundant nor obviously larger than those produced in summer. Because the wood of an annual ring has vessels located throughout it, it is said to be diffuse porous (Fig. 7c), whereas species with vessels restricted mainly to early wood are ring porous. Examples of trees with diffuse porous wood are yellow birch, aspen, sugar maple, and American holly; trees with ring porous wood include red oak, sassafras, and honey locust. In mild tropical climates, the cambium may remain active almost continuously, and the wood of one year is difficult to distinguish from that of another; annual rings are indistinct.

FIGURE 7: (a) In this transverse section, portions of five annual rings are visible. The lowermost ring is oldest, the next higher ring was formed 1 year after the lowest ring, and so forth. The vascular cambium is located beyond the top of the photograph. The wood is ring porous, and vessel-rich early wood is easily distinguished from fiber-rich late wood (transverse section; X 50). (b) This wood is very strongly ring porous, with large vessels formed only when the cambium first becomes active. Later, the cambium produces fibers exclusively, except for a few rare, very narrow vessels. Two large multiseriate rays are visible, but most rays are narrow and uniseriate (transverse section; X 15). (c) This wood is diffuse porous, with vessels occurring rather uniformly in both late wood as well as early wood. The annual rings are not as conspicuous as in ring porous wood (transverse section; X 40). (d) Most gymnosperms have wood composed only of tracheids in their axial system, but they still have early wood (wide tracheids) and late wood (narrow tracheids) (transverse section; X 50).
HEARTWOOD AND SAPWOOD
The center of a log is almost always darker in color than the outer wood, and it is usually drier and more fragrant (Fig. 8). The dark wood is heartwood and the lighter, moister outer region is sapwood. The different regions exist because vessels and tracheids do not function forever in water conduction; water columns break due to freezing, wind vibration, tension, wood-boring insects, and other factors. Once the water column breaks, the upper portion of the water column is drawn upward to the leaves and the lower portion falls toward the roots. After contact with the upper portion is broken, there is no means of pulling the water in the lower portion upward; vessels and tracheids in which this has occurred usually never conduct water again. Although only a few water columns break at any time, all water columns in an annual ring eventually snap. The conducting capacity of the wood is decreased, but more tracheary elements are produced by the cambium during the next year. However, a vessel is wide enough that a fungus can easily grow up through it; for tracheary elements that are not conducting, a mechanism that seals them off is selectively advantageous. The adjacent wood parenchyma cells push bubbles of protoplasm through the pits into a vessel, forming a plug, called a tylosis (plural: tyloses), completely across it (Fig. 9). This occurs repeatedly, and the vessel may become filled. These and other parenchyma cells, both axial and ray parenchyma, undergo numerous metabolic changes and produce large quantities of phenolic compounds, lignin, and aromatic substances that inhibit growth of bacteria and fungi. Ultimately, the parenchyma cells die. This heartwood is aromatic and decay resistant.

FIGURE 8: The vascular cambium was almost perfectly circular in cross-section in this tree trunk. Note that the innermost annual rings, formed when the tree was a sapling, are wider than the outer rings—the tree grew more vigorously when it was younger. The dark region is heartwood and the narrow light region is sapwood; if the tree had lived a few years longer, several more of the innermost rings of sapwood would have converted to heartwood. Bark-secondary phloem-is also present. (William E. Ferguson).

FIGURE 9 :(a) Tyloses form as protoplasm from surrounding parenchyma cells pushes into the vessel. This was just beginning to form tyloses when the wood was collected for microscopy. Boiling destroyed the protoplasm (transverse section; X 250). (b) The vessels of this wood are completely occluded by tyloses; this sample came from a piece of heartwood (X 50).
The outer, younger region, the sapwood, contains living parenchyma as well as water- filled tracheary elements; it is full of "xylem sap"—hence its name. It is gradually converted to inactive wood as its water columns break; then as the parenchyma cells become resistant to fungi and die, it is converted to heartwood. A new layer of sapwood is formed each year by the vascular cambium, and on average one annual ring is converted to heartwood each year. Thus, whereas the heartwood becomes wider with age, the sapwood has a more or less constant thickness (Table 2). Of course this is not true of young stems and roots: Black walnut typically has wood that functions for 10 years before converting to heart- wood. In a seedling or a branch only 9 years old, no heartwood is present yet.

In branches or trunks that are not vertical, gravity causes a lateral stress; if not counteracted, the branch would droop and become pendant. In response to such stress, most plants produce reaction wood (Fig. 10). In dicots, this develops mostly on the upper side of the branch and is known as tension wood. In a cross-section of such a branch, the growth rings are eccentric, being much wider on the top of the branch. Tension wood contains many special gelatinous fibers whose walls are rich in cellulose but have little or no lignin. These fibers exert tension on the branch, preventing it from drooping, or the tension wood may even contract, slowly lifting a branch to a more vertical orientation. Conifers form reaction wood that is located on the underside of the branch and is known as compression wood. It is enriched in lignin and has less cellulose; the growth rings are especially wide on the lower side of the limb.

FIGURE 10 :Reaction wood in poplar (Populus). These are called gelatinous fibers because they have a soft inner layer that is deformed when specimens are cut. Reaction wood is rich in gelatinous fibers (X 600). (Courtesy of W. A. Cote, State University of New York).



|
|
|
|
دراسة يابانية لتقليل مخاطر أمراض المواليد منخفضي الوزن
|
|
|
|
|
|
|
اكتشاف أكبر مرجان في العالم قبالة سواحل جزر سليمان
|
|
|
|
|
|
|
اتحاد كليات الطب الملكية البريطانية يشيد بالمستوى العلمي لطلبة جامعة العميد وبيئتها التعليمية
|
|
|