
آخر المواضيع المضافة

 النبات
الحيوان
الأحياء المجهرية
علم الأمراض
التقانة الإحيائية
التقنية الحيوية المكروبية
التقنية الحياتية النانوية
علم الأجنة
الأحياء الجزيئي
علم وظائف الأعضاء
الغدد
المضادات الحيوية
النبات
الحيوان
الأحياء المجهرية
علم الأمراض
التقانة الإحيائية
التقنية الحيوية المكروبية
التقنية الحياتية النانوية
علم الأجنة
الأحياء الجزيئي
علم وظائف الأعضاء
الغدد
المضادات الحيوية| Introduction to Genetics |


|
|
Read More
Date: 5-11-2015
Date: 5-11-2015
Date: 2-11-2015
|
Introduction to Genetics
Genetics is the study of inheritance or heredity. It deals with the transmission of characters, specific for that particular type of plant or animal, from the parent to the offspring of the next generation.
The history of Genetics is closely linked with the ancient cultural his-tory of man. It began ten thousand years ago, when human beings changed from a nomadic life to the life of settlers. As settlers in early civilization they started living as groups. A need for food forced them to explore nature and adopt agriculture. The agricultural revolution in early human civilization opened the doors for very rapid improvements in human culture. They learnt to identify plants that could be grown and cultivated. Several cultivation processes evolved. They started selecting suitable varieties for agriculture.
Simultaneously they started domestication of animals. Many animals were brought under human control. Gradually they understood their reproductive methods. Hybridization and generation of new varieties of animals like horses, dogs, cats and cattle happened. The idea of Genetics was adopted in everyday practice.
A scientific approach to understand inheritance was initially made by Gregor Mendel. He did hybridization experiments in Pisum sativum more out of curiosity and his devotion to science. He applied statistical methods to biological research.
Since the ‘rediscovery’ of Mendelism in 1900, Genetics has made rapid developments. The advancement in cell biology, microscopy, biochemistry and other disciplines provided the necessary infrastructure for this science to reach its climax. As of to-day all processes and causes for inheritance are well understood. Through the developments in Genetic Engineering, Biotechnology, Tissue culture and other methods we are trying to exploit nature. However, the steady processes of dominance of human beings on earth through their knowledge of science and skill in technology will also ensure the safety of earth in the times to come. In such a safeguarding the science of Genetics with all its related fields will play a key role.
: Multiple Alleles
According to Mendelism, a genetical character is controlled by the combined action of two genes or factors in the same loci of two homologues chromosomes. It means that each gene has two alternative forms or alleles and their expression are known as allelomorphs. Of these two, one is dominant and the other recessive. However, some genetical characters are determined by several forms of an allele known as multiple alleles. There are many examples for multiple allelism.
1-Skin colour in rabbits
Rabbits have five kinds of skin colour, coloured (agouti), chinchilla, himalayan albino and albino, light grey. Of these, the chinchilla variety is lighter in colour. Himalayan albino will have pink eyes, white coat colour and black colour in tips like nose, tail and feet.

A crossing of a homozygous coloured rabbit with an albino resulted in following F1 and F2 generation.
Parents CC x ca ca
(coloured) (albino)
F1 Cca x Cca
(coloured)
F2 CC Cca caca
25% 50% 25%
(coloured) (coloured) (albino)
This result shows that coloured condition is dominant over albino.
Other possible crossings:
Parents CC x ch ch
(coloured) (Himalayan albino)
F1 Cch x Cch
(coloured)
F2 CC Cch chch
25% 50% %25
( (coloured) (coloured) (Himalayan albino
Parents chch x ca ca
(Himalayan albino) (albino)
F1 chca x chca
(Himalayan albino)
F2 chch chca caca
25% 50% %25
Himalayan albinos Himalayan albinos albino
Parents cchcch x caca
(chinchilla) (albino)
F1 cch ca x cch ca
(light grey)
F2 cchcch cchca caca
25% 50% 25
(chinchilla) (light grey) (albino)
In chinchilla, coat colour is lighter than the coloured (agouti). The coloured character is dominant over chinchilla. However F1 hybrids between chinchilla and himalayan albino (Cchch) or between chinchilla and albino (cchca) show light grey skin colour. The occurrence of light grey colour is due to partial expression of the gene for chinchilla in a heterozygous state.
Genotype Phenotype
CC, Ccch, Cca ,Cch coloured (wild)
cchcch chinchilla
cch ch ,cchca light grey
chch ,chca himalayan albino
caca albino
2-ABO blood groups in human beings
The ABO blood group system in human beings was established by K. Landsteiner. It is based on the presence or absence of certain antigens. There can be two antigens A or B in the blood, resulting in four blood groups, namely A, B, AB and O. These are called ABO blood groups or Landsteiner blood groups.
The inheritance of ABO system illustrates a new principle in genetic control of phenotypes.
The blood of a person having a group will have the antigen A and a person having B group will have the antigen B. With these antigens A and B there are certain naturally occurring antibodies in the serum of the blood. The antibodies in a particular individual will be found only for those antigens which are absent in blood of this individual. The presence of antigens and antibodies
Occur as follows
Blood groups Antigen Antibody in the serum
A A anti B
B B anti A
AB A and B None
O None anti A and B
Antibodies in the blood of ‘group A’ will agglutinate red blood corpuscles of the blood group B. Similarly the antibody in blood ‘group B’ will agglutinate red blood corpuscles of the blood group A. Since no antibody is found in ‘group AB’ blood, it will not agglutinate any other group. ‘Group O’ will have antibodies for ‘group A’ and B. Hence ‘group O’ will agglutinate group A and B.
Compatibility of donor blood to that of the recipient will occur as follows.
Blood group of the donor Blood group of the recipient
A A and AB
B B and AB
AB AB
O O, A, B, AB
From the table provided it is obvious that ‘group AB’ is universal recipient. ‘Group O’ is universal donor.
The gene for ABO system is conventionally represented by the symbol ‘I’. Alleles IA and IB code for the enzymes involved in the formation of the antigens A and B respectively and the allele ‘Io’ for no known protein. Thus the genes can result in 6 possible genotypes but four possible phenotypes as found in the table
Blood group possible genotype
O IoIo
A IAIA or IAIO
B IBIB or IBIO
AB IAIB
The alleles IA and IB jointly express themselves in the individual, they are codminant. Both IA and IB are dominant to the recessive allele Io.
Disputed parentage and blood groups
The identification of blood group may help to decide in cases concerned with parentage issues. By knowing the blood groups of parents, it is possible to determine the possible blood groups in the children. The impossibility of a particular blood group in the progeny can also be pointed out.
Parents Blood group in progeny Impossible blood groups
O x O O A, B, AB
O x A O, A B, AB
O x B O, B A, AB
O x AB A, B O, AB
A x A A, O B, AB
A x B A, B,AB, O None
B x B B, O A, AB
A x AB A, B, AB O
B x AB A, B, AB O
AB x AB A, B, AB O
Rh blood group
Rh factor in blood was discovered by Landsteiner and Wiener in 1940. It was initially discovered in rabbits, immunized with the blood of Rhesus monkey. The human beings whose blood will get agglutinated with rabbit serum were designated as Rh+, and whose blood if not agglutinated were designated as Rh-. Wrong transfusion can cause agglutination of blood in the recipient hence before transfusion of blood, along with identification of ABO blood group, it is necessary to test compatibility of Rh factor.
The presence of Rh+ child in the uterus of the Rh- mother can cause agglutination in the blood of the fetus. Even though such an unfortunate incident may not happen in the first pregnancy, it could occur in successive pregnancies. The death of the foetus in such cases is due to hemolytic anemia. This disease is called erythroblastosis fetalis.
Quantitive inheritance :
Charles Darwin while working on his theory of Natural selection recognized two principal types of variations, namely continuous and discontinuous. The continuous variations show the whole range of variations in a particular character. The discontinuous variations would appear all of a sudden and show no gradation.
Mendel in his work depended on sharp or alternate characters comprising discontinuous variations. For example, when tall and dwarf plants were crossed, in F1 only tall plants appeared. Crossing F1 plants, produced only tall and dwarf plants in the F2 generation. Mendel discarded intermediate characters if any, seen.
However, by the end of the 19th century Galton, a British geneticist and statistician was interested in the study of continuous variations. He tried to find an answer for its origin. He called these characters as metrical characters and found them inherited.
Thus in the beginning of the 20th century two groups of geneticists emerged. They were the mendelians and the biometricians. While the mendelians considered that all hereditary differences are discontinuous and qualitative, the biometricians believed that hereditary variations are basically continuous and quantitative. These two views remained contradictory. Later, Johansen (1903), through his work on bean seeds proved that both the views of mendelians and biometricians were only partly correct.
Yule (1906) suggested that quantitative variations may be controlled by large number of individual genes, with each gene having a small effect. Later on, such genes were called as polygenic systems. The hereditary processes operating through such system was explained through multiple factor hypothesis.
Multiple factors
Skin colour in human beings
The inheritance of skin colour in black and white populations in United States of America was worked out and reported by C. B. Davenport in 1913. In USA, marriages between black and white individuals has resulted in a population known as mulattoes. They have intermediate skin colour in the first generation. When the mulattoes marry among themselves, all shades of skin colour are obtained.
If the skin colour is due to genes in two loci A and B, the genotype of black and white persons will be AABB and aabb respectively. The genotypes of mulattoes will be AaBb. However the F2 generation has yielded five or more shades of skin colour. The observed results on number of individuals with differing shades indicate the involvement of four or five gene pairs in the control of skin colour. The effect of these genes may be further modified due to modifying genes.
Studies have also shown that, the control of most characteristics is multifactorial. For example, in human beings several different gene loci are involved in determining characteristics such as body height and body mass.
An understanding of the polygenic influence on a specific genetic trait requires enormous data. The data are to be analyzed employing several bio- statistical and bio-mathematical tools.
From such studies it becomes clear that while at individual level the basic inheritance remains mendelian, at the population level it appears much more complicated.
Sex Determination :
Differentiation as male and female sex and reproduction through sexual processes are basic phenomena in the living world. Sexual reproduction is a strategy for the production of diversity in nature. It is achieved through efficient functional anatomy of male and female individuals. In this natural system, the sex cells and the basic reproductive organs are the primary sexual characters. Morphological, Physiological and behavioral characters exhibited by male and female of a species are called the secondary sexual characters. Thus the characteristic differences between male and female animals constitutes sexual dimorphism.
The fundamental mechanism concerned with sex determination is genetical in nature. These genetical methods vary in different animals.
A. Sex Chromosome mechanism
The chromosomal basis for the determination of sex was first proposed by Clarance Mc clung in 1902. He observed gametogenesis in grasshopper (Xiphidium fasciatum).He reported that while the somatic cells of the females had 24 chromosomes the males had 23 chromosomes. Similar observations were made in bugs and beetles by several other workers.
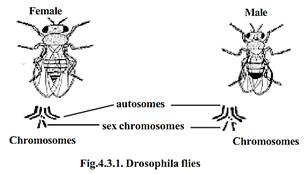
By further studies it was concluded that in dioecious organisms there are two types of chromosomes. Among them the chromosomes concerned with body or somatic characters were named as autosomes (A). The other type of chromosomes concerned with sex determination were called sex chromosomes (X and Y).
Later it was found that sex chromosomes (X and Y) had structural differences. The cytological studies have shown that X-chromosomes in most cases are straight, rod-like and slightly longer. The Y-chromosome was found to be smaller with one end curved or bent to one side (as in Drosophila).

Based on sex chromosomal determination of sex, there may be two types of organisms, namely 1. Heterogametic males 2. Heterogametic females.
Heterogametic males: - In this type of sex determination the female has two X-chromosomes. The males have only one X-chromosome. During gametogenesis the males could produce two types of gametes i.e., one type having ‘X-chromosomes’ and the other without ‘X-chromosome’. Hence the males could be called the heterogametic sex. The females can produce only one type of gametes i.e., all eggs having one ‘X’-chromosome each. Hence the female could be called the homogametic sex. Each type of sex determination has subtypes as shown in the table.
B. Genic balance mechanism
Further studies on sex determination showed that sex determination was not the inheritance of genes by the sex chromosomes alone. Studies on intersex and supersex indicated the operation of still more a complex mechanism for the determination of sex. Thus the genic balance mechanism of sex was discovered. It was first described by C.B Bridges in 1921.
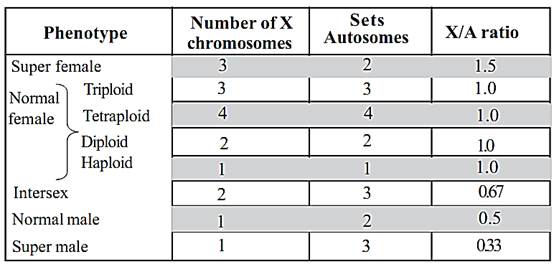
In Drosophila it was discovered that the sex of an individual depends upon the ratio of x chromosomes to the autosomes. For sex determination, each haploid set of autosomes carry factors with a male determining value equal to one (1). Each x chromosome carries factors with a female determining value of one and a half (1 .5 ). Hence in a normal male (AAXY), the male female determinants are in the ratio of 2 : 1.5(‘A’ represents a haploid set of autosomes). Thus the genic balance is in favour of maleness. A normal female (AAXX) has the male female determination ratio of 2:3. Therefore the balance is in favour of femaleness.
Sex determination in Human beings
The human sex determination mechanism to a larger extent re-sembles XX - XY type of Drosophila. However, the Y chromosome contains male determining genes and it is the determiner of fertility and sex of male individual.
Thus in human beings, the presence of Y chromosome deter-mines maleness and its absence determines femaleness. Evidences to estab¬lish this viewpoint had been provided by certain abnormal conditions called syndromes.
Sex anomalies in Human beings
1-Turner’s syndrome (XO Females): In this abnormality the females are sterile and have short stature. They have webbed neck, broad shield-shaped chest, low intelligence, under developed breasts and poorly developed ovaries. These conditions result due to the presence of 44 autosomes and only one X chromosome in her body cells. This abnormality is known as Turner’s syndrome.
2-Klinefelter’s syndrome: - This syndrome is caused due to the presence of an extra X chromosome in males. This happens when an abnormal egg with XX chromosome is fertilized by a sperm carrying Y chromosome. The zygote will have three sex chromosomes (XXY). The resulting young one is an abnormal sterile male. The symptoms of this syndrome are the presence of small testicles, mental retardation, longer arms and high pitched voice.
3-Super females: - These females are also known as Poly X females. They possess an extra X-chromosome (44 autosomes +3 x chromosomes). The poly X females are mentally retarded and sterile
4-XYY males: - Such males will have an extra Y chromosome (XYY). This condition results in mental retardation and criminal.
5-Hermaphroditism :- A hermaphroditic person will have one extra X and Y chromosome. The person will have both ovarian and testicular tissues. The external genitalia will not be well defined.
C. Male haploidy or Haplo-diploidy mechanism
This mechanism is also known as arrhenotokus parthenogenesis. It is a common mechanism in several insects such as ants, bees and wasps. In these insects, fertilized eggs develop into diploid females and unfertilized egg into haploid males.
In a honey bee colony a queen bee can lay two types of eggs. They are the fertilized and unfertilized eggs. It happens by controlling the sphincter in the sperm receptacle of the female. The diploid female zygote can develop either into a queen or a sterile female worker bee. The sterile nature of the worker bee is due to poor nourishment. The haploid zygote develops into a male. This mechanism of sex determination helps to maintain the polymorphic nature in a honey bee colony.
Apart from genetical systems, the sexuality can also be controlled by various factors such as metabolism, environment and hormones.
Sex in Bonellia
Bonellia viridis is a marine worm. Its sex determination was studied by F. Baltzer(1935). The adult female worm is about 2.5 cm long. It has a well-defined anatomical organization.
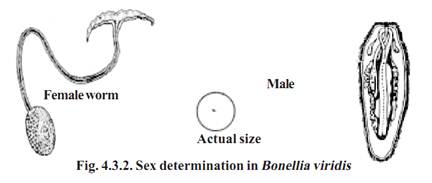
The male is very small and microscopic. Its body organs are rudimentary. The males normally live as parasites attached to females. All larvae of Bonellia are genetically similar. However a larva settling on the proboscis of an adult female becomes a male individual. If a larva develops in isolation (i.e., in the absence of a female) it develops into female. If a developing male is detached from the proboscis of female, it becomes an intersex. From these observations it could be inferred that the proboscis of adult female secretes some hormone like substance and that substance suppresses femaleness and induces maleness in the larvae which remains attached.
Sex - linked Inheritance
Most of the inheritable characters are controlled by genes located in autosomes. The inheritance of traits related to autosomes normally follows Mendel’s laws. Mendelian ratios are not obtained for those characters for which genes are exclusively located either in X or Y chromosome. The genes that occur only on X chromosomes are called as X - linked genes. Similarly, that the genes occur exclusively on Y chromosomes are called the holandric genes. The inheritance of X or Y linked genes is called as sex linked inheritance. Thus the sex linked inheritance may be X- linked, Y- linked or XY linked.
X - Linked inheritance
T. H. Morgan (1910) in his studies on inheritance of genes in Drosophila discovered that the patterns of inheritance of certain traits were found to vary with the sex of the parent and offspring. He found that the gene white eye colour is X - linked. Further it was found to be recessive to another X - linked, dominant gene for red eye colour.
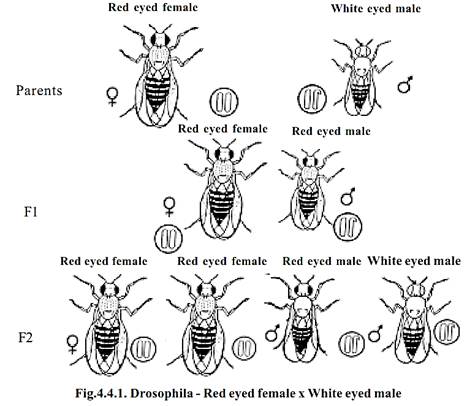
Red eyed female x white eyed male
When a wild red eyed female Drosophila is crossed with a mutant white eyed male, all the F1 individuals (males and females) have red eyes. When the red eyed male and red eyed female of the F1 were intercrossed, in the F2 generation all the female flies were found to be red eyed. Among the males 50 % had red eyes and another 50 % had white eyes.
White eyed female x Red eyed male
When a white eyed female Drosophila is crossed with a red eyed male, all the female individuals in the F1 generation are red eyed and all the males are white eyed. When these red eyed female individuals and white eyed male individuals of F1 are intercrossed the F2 generation possessed 50 % red eyed and 50 % white eyed females. Similarly the male population of F2 included 50 % red eyed and 50 % white eyed flies.
Sex linked inheritance in Humans
Most of the sex linked characters in humans are X - linked. There are 150 confirmed X- linked traits known. Most of them are recessives.
Colour blindness:
The human vision is basically due to cells called rods and cones found on the retina of the eye. The cone cells are sensitive to red, green and violet light. The formation of colour sensitive cones is controlled by a dominant X- linked gene.
The recessive form of this gene is incapable of producing colour sensitive cones. Hence homozygous recessive females (XCXC) and Hemizygous recessive males (XCY) are unable to differentiate between red and green colour. The frequency of colour blind women is less than colour blind men.
Colour - blind man x normal visioned woman
When a colour-blind man marries a normal woman in their F1 progeny all children would be normal. However the female will be a carrier for the recessive gene. If that female gets married to a normal male in the F2 generation normal and colour-blind nature will occur in 3: 1 ratio.
Pleiotropy
It is an established fact that a specific gene controls a specific phenotypic trait. This finding is not always true. Studies on ‘gene expression’ have revealed that a gene often influences more than one phenotypic trait. This phenomenon of multiple effects of a single gene is called pleiotropism. In such a genic influence more conspicuous expression of a phenotypic trait by a gene is called its major effect. If the gene causes other less conspicuous phenotypic changes, it is known as secondary effect. Such genes responsible for multiple effects are called pleiotropic genes.
Vestigieal wings in Drosophila are caused by a recessive gene in homozygous condition. A keen observation has shown that this gene affects other traits as well. They are
i) The small halters or balancers behind the wing
ii) structure of reproductive organs
iii) Egg production
iv) Life duration and
v) Bristles on the body
References
T. Sargunam Stephen, Biology (Zoology). First Edition – 2005, Government of Tamilnadu



|
|
|
|
علامات بسيطة في جسدك قد تنذر بمرض "قاتل"
|
|
|
|
|
|
|
أول صور ثلاثية الأبعاد للغدة الزعترية البشرية
|
|
|
|
|
|
|
مكتبة أمّ البنين النسويّة تصدر العدد 212 من مجلّة رياض الزهراء (عليها السلام)
|
|
|
















