
آخر المواضيع المضافة

 النبات
الحيوان
الأحياء المجهرية
علم الأمراض
التقانة الإحيائية
التقنية الحيوية المكروبية
التقنية الحياتية النانوية
علم الأجنة
الأحياء الجزيئي
علم وظائف الأعضاء
الغدد
المضادات الحيوية
النبات
الحيوان
الأحياء المجهرية
علم الأمراض
التقانة الإحيائية
التقنية الحيوية المكروبية
التقنية الحياتية النانوية
علم الأجنة
الأحياء الجزيئي
علم وظائف الأعضاء
الغدد
المضادات الحيوية| The Holoenzyme Goes Through Transitions in the Process of Recognizing and Escaping from Promoters |


|
|
Read More
Date: 28-10-2020
Date: 19-5-2016
Date: 9-12-2015
|
The Holoenzyme Goes Through Transitions in the Process of Recognizing and Escaping from Promoters
KEY CONCEPTS
- When RNA polymerase binds to a promoter, it separates the DNA strands to form a transcription bubble and incorporates nucleotides into RNA.
- A cycle of abortive initiations may occur before the enzyme moves to the next phase.
- Sigma factor is usually released from RNA polymerase when the nascent RNA chain reaches approximately 10 bases in length.
We can now describe the stages of transcription in terms of the interactions between different forms of RNA polymerase and the DNA template. The initiation reaction can be described by the parameters that are summarized in FIGURE 1:
- The holoenzyme–promoter reaction starts by forming a closed binary complex, as shown in Figure 1a. “Closed” means that the DNA remains duplex. The formation of the closed binary complex is reversible; thus, it is usually described by an equilibrium constant (KB ). The values of the equilibrium constant range widely for forming the closed sequence-dependent complex.
- The closed complex is converted into an open complex of 1.3 turns of the double helix in a series of steps by first “melting” a short region of DNA around the −10 region, giving an unstableintermediate open complex within the sequence bound by the enzyme, as shown in Figure 2b. For most promoters, conversion from the closed to the open complex is irreversible, and this reaction can be described by the forward rate constant (kf). Some promoters (e.g., rRNA promoters), though, do not form stable open complexes, and this is a key to their regulation. Sigma factor plays an essential role in the melting reaction (see the sections later in this chapter on sigma factors). The transitions that occur from initiation to elongation are also accompanied by major changes in the structure and composition of the complex.
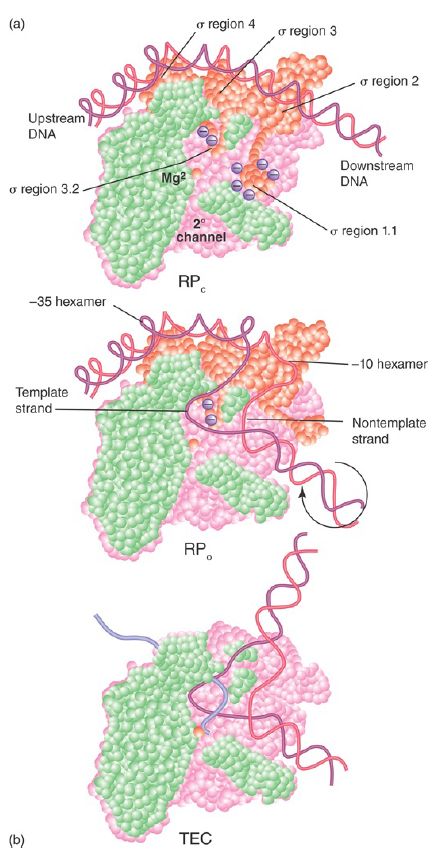
FIGURE 1. RNA polymerase passes through several steps prior to elongation. A closed binary complex is converted to an open form and then into a ternary complex.
Data from S. P. Haugen, W. Ross, and R. L. Gourse, Nat. Rev. Microbiol. 6 (2008): 507– 519.
Changes in the shape of RNA polymerase accompany the kinetic transitions described earlier, as well as the transition to the elongation complex (as illustrated in FIGURE 2). In the closed complex, RNA polymerase holoenzyme covers about 55 bp of DNA, extending from about −55 to about +1. The double-stranded DNA binds primarily along one face of the holoenzyme, contacting the Cterminal domains of the α subunits as well as regions 2 and 4 of the σ subunit (see Figure 2). During the transition to the open complex, the conformation of both the RNA polymerase and the DNA change. The most dramatic changes in the structure of the complex are depicted in Figure 1: (1) an approximately 90° bend in the DNA, which allows the template strand to approach the active site of the enzyme; (2) strand opening of the promoter DNA between about −11 and +3 with respect to the transcription start site; (3) scrunching of the promoter DNA into the active channel, forming the transcription bubble; and (4) closing of the jaws of the enzyme to encircle the section of the promoter downstream of the transcription start site. Thus, promoter contacts in the open complex extend from about −55 to about +20.
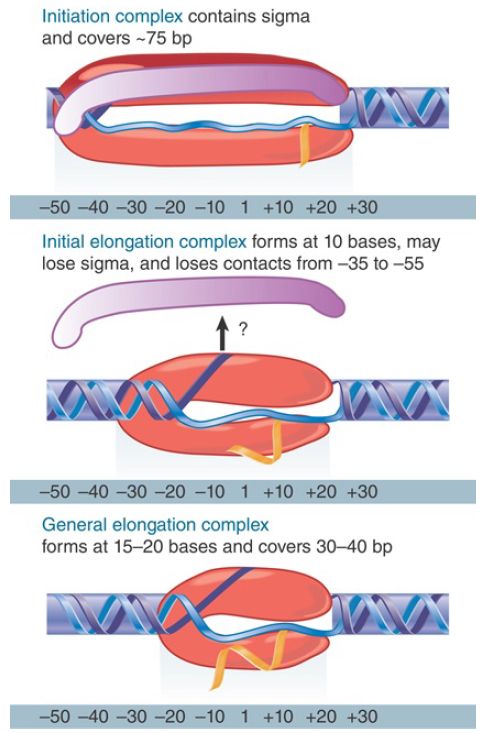
FIGURE 2. RNA polymerase initially contacts the region from −55 to +20. When sigma dissociates, the core enzyme contracts to −30; when the enzyme moves a few base pairs, it becomes more compactly organized into the general elongation complex.
The next step is to incorporate the first two nucleotides and to form a phosphodiester bond between them. This generates a ternary complex containing RNA as well as DNA and the enzyme. At most promoters, an RNA chain forms that is several bases long without movement of the enzyme down the template. After each base is added, there is a certain probability that the enzyme will release the RNA chain, resulting in abortive initiation products. After release of the abortive product, the enzyme again begins synthesizing RNA at position +1. Repeated cycles of abortive initiation generate oligonucleotides that usually are only a few bases long, but that can be almost 20 nucleotides in length, before the enzyme actually succeeds in escaping from the promoter.
Interactions with RNA polymerase ultimately dissolve during the process of promoter escape. By the time the RNA chain has been extended to 15 to 20 nucleotides, the enzyme generally has gone through all the transitions that typify an elongation complex. The two most obvious of these transitions are the release of the sigma factor, shown in Figure 2, and the formation of a complex covering only about 35 bp of DNA, rather than the approximately 70 bp characteristic of promoter complexes. Although release of sigma factor usually occurs during the process of promoter escape, this is not obligatory for the transition to elongation. In some cases sigma factor has been identified in elongation complexes, but its association with the enzyme may reflect rebinding to the core enzyme during the elongation phase.



|
|
|
|
علامات بسيطة في جسدك قد تنذر بمرض "قاتل"
|
|
|
|
|
|
|
أول صور ثلاثية الأبعاد للغدة الزعترية البشرية
|
|
|
|
|
|
|
مدرسة دار العلم.. صرح علميّ متميز في كربلاء لنشر علوم أهل البيت (عليهم السلام)
|
|
|
















