
آخر المواضيع المضافة

 النبات
الحيوان
الأحياء المجهرية
علم الأمراض
التقانة الإحيائية
التقنية الحيوية المكروبية
التقنية الحياتية النانوية
علم الأجنة
الأحياء الجزيئي
علم وظائف الأعضاء
الغدد
المضادات الحيوية
النبات
الحيوان
الأحياء المجهرية
علم الأمراض
التقانة الإحيائية
التقنية الحيوية المكروبية
التقنية الحياتية النانوية
علم الأجنة
الأحياء الجزيئي
علم وظائف الأعضاء
الغدد
المضادات الحيوية| Transposons Cause Rearrangement of DNA |


|
|
Read More
Date: 7-3-2021
Date: 3-12-2015
Date: 21-12-2015
|
Transposons Cause Rearrangement of DNA
KEY CONCEPTS
- Homologous recombination between multiple copies of a transposon causes rearrangement of host DNA.
- Homologous recombination between the repeats of a transposon may lead to precise or imprecise excision.
In addition to the “simple” intermolecular transposition that results in insertion at a new site, transposons promote other types of DNA rearrangements. Some of these events are consequences of the relationship between the multiple copies of the transposon. Others represent alternative outcomes of the transposition mechanism, and they leave clues about the nature of the underlying events.
Rearrangements of host DNA may result when a transposon inserts a copy at a second site near its original location. Host systems may undertake reciprocal recombination between the two copies of the transposon; the consequences are determined by whether the repeats are in direct or inverted orientation.
FIGURE 1. illustrates the general rule that recombination between any pair of direct repeats will delete the material between them. The intervening region is excised as a circle of DNA (which is lost from the cell); the chromosome retains a single copy of the direct repeat. A recombination between the directly repeated IS1 modules of the composite transposon Tn9 would replace the transposon with a single IS1 module.
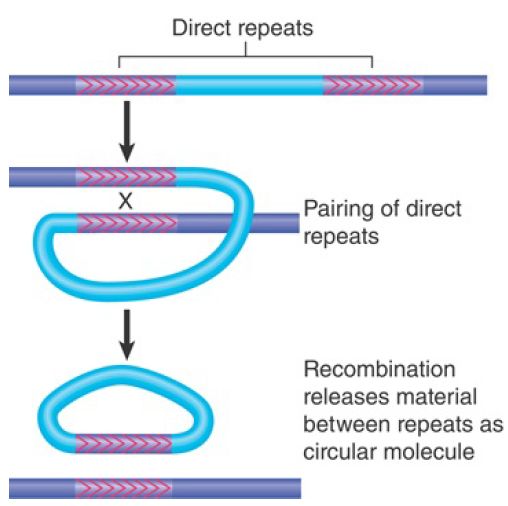
FIGURE 1. Reciprocal recombination between direct repeats excises the material between them; each product of recombination has one copy of the direct repeat.
Deletion of sequences adjacent to a transposon could therefore result from a two-stage process; transposition generates a direct repeat of a transposon, and recombination occurs between the repeats. The majority of deletions that arise in the vicinity of transposons, however, probably result from a variation in the pathway followed in the transposition event itself.
FIGURE 2 depicts the consequences of a reciprocal recombination between a pair of inverted repeats. The region between the repeats becomes inverted; the repeats themselves remain available to sponsor further inversions. A composite transposon whose modules are inverted is a stable component of the genome, although the direction of the central region with regard to the modules could be inverted by recombination.
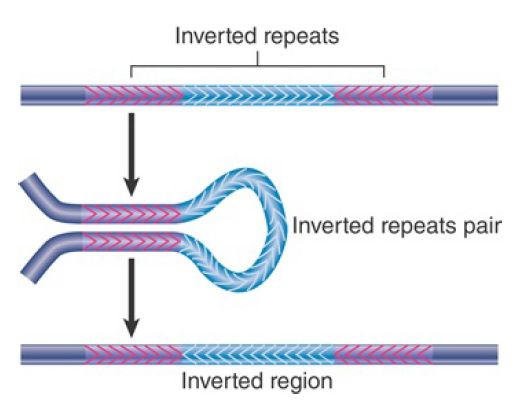
FIGURE 2. Reciprocal recombination between inverted repeats inverts the region between them.
Excision in this case is not supported by transposons themselves, but occurs when bacterial enzymes recognize homologous regions in the transposons. This is important because the loss of a transposon may restore function at the site of insertion. Precise excision requires removal of the transposon, plus one copy of the duplicated sequence. This is rare; it occurs at a frequency of approximately 10–6 for Tn5 and 10–9 for Tn10. It probably involves a recombination between the duplicated target sites.
Imprecise excision leaves a remnant of the transposon. The remnant may be sufficient to prevent reactivation of the target gene, but it may be insufficient to cause polar effects in adjacent genes so that a change of phenotype occurs. Imprecise excision occurs at a frequency of 10–6 for Tn10. It involves recombination between sequences of 24 bp in the IS10 modules; these sequences are inverted repeats, but because the IS10 modules themselves are inverted, they form direct repeats in Tn10.
The greater frequency of imprecise excision compared with precise excision probably reflects the increase in the length of the direct repeats (24 bp as opposed to 9 bp). Neither type of excision relies on transposon-encoded functions, but the mechanism is not known.
Excision is RecA independent and could occur by some cellular mechanism that generates spontaneous deletions between closely spaced repeated sequences.
Both precise and imprecise excisions can also arise as a consequence of transposition of cut-and-paste elements in eukaryotes. In this case, the outcome depends on the nature of the repair of the double-stranded DNA break introduced by excision of the element. This break can be repaired using the homologous chromosome or the sister chromatid, resulting in a transfer of DNA from those templates. Repair using a chromosome that lacks the transposon insertion can result in precise restoration of sequences surrounding the original insertion. Repair using the sister chromatid results in restoration of the transposon insertion. Incomplete repair can result in deletions, either of sequences flanking the insertion or of portions of the transposon. Alternatively, the break can be repaired using nonhomologous end joining, which results in the addition or deletion of short stretches of DNA.



|
|
|
|
علامات بسيطة في جسدك قد تنذر بمرض "قاتل"
|
|
|
|
|
|
|
أول صور ثلاثية الأبعاد للغدة الزعترية البشرية
|
|
|
|
|
|
|
مكتبة أمّ البنين النسويّة تصدر العدد 212 من مجلّة رياض الزهراء (عليها السلام)
|
|
|
















