
آخر المواضيع المضافة

 النبات
الحيوان
الأحياء المجهرية
علم الأمراض
التقانة الإحيائية
التقنية الحيوية المكروبية
التقنية الحياتية النانوية
علم الأجنة
الأحياء الجزيئي
علم وظائف الأعضاء
الغدد
المضادات الحيوية
النبات
الحيوان
الأحياء المجهرية
علم الأمراض
التقانة الإحيائية
التقنية الحيوية المكروبية
التقنية الحياتية النانوية
علم الأجنة
الأحياء الجزيئي
علم وظائف الأعضاء
الغدد
المضادات الحيوية| Class Liliopsida |


|
|
Read More
Date: 30-10-2015
Date: 23-11-2016
Date: 27-11-2016
|
Class Liliopsida
At present, monocots are widely believed to have arisen from early dicots about 80 to 100 million, perhaps even 120 million, years ago. Certain features common to most monocots probably are inherited from their dicot ancestors. Because all monocots lack ordinary secondary growth and wood, the dicot ancestors were probably herbs with either no vascular cambium or little cambial activity. The gynoecia of many monocots are composed of several carpels that are either free of each other or at most only slightly fused together. The perianth is composed of sepals and petals that are usually relatively unspecialized and have very little fusion. These features must have been present in the group from which the monocots evolved. The dicot order Nymphaeales, which contains water lilies, has similar features. An early set of dicots may have resembled Nymphaeales and may have given rise to both the water lilies and the monocots (Fig. 1).

FIGURE 1:The flowers of water lilies have numerous sepals, stamens, and so on, with all parts arranged in spirals like the leaves of a rosette plant. This is believed to be a relictual feature similar to a gymnosperm cone. Monocot flowers would be derived from an ancestral flower like this by mutations that result in only three or six of each type of floral appendage.
One theory about the parallel venation of monocot leaves postulates that the ancestors had broad leaves and lived as aquatic plants. Gradually the leaves evolved to a more reduced, simple type without a blade, a form that is more adaptive for a submerged leaf (Fig. 2a and b). Some of these plants moved into drier habitats where their leaves were not submerged and a broad or long lamina would be advantageous. Mutations that resulted in a basal meristem were selected, resulting in strap-shaped leaves (Fig. 2c). In the evolutionary lines of "broadleaf" monocots such as palms, philodendrons, and dieffenbachias, a marginal type of meristem evolved; it was located at the end of the residual leaf, resulting in the formation of what appears to be a petiole and lamina (Figs. 2 d and 11).
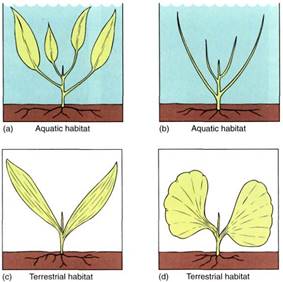
FIGURE 2: (a) Monocots are believed to have originated as semiaquatic plants that inhabited swamps and marshes. (b) The leaf may have consisted only of a leaf base and petiole, the entire blade having been lost evolutionarily. (c) A "pseudolamina" evolving from a petiole. (d) A "broad leaf" evolving from a petiole.
Out of these first monocots, five subclasses evolved: (1) Alismatidae, (2) Arecidae, (3) Commelinidae, (4) Zingiberidae, and (5) Liliidae (Fig. 3). We do not know which group evolved first; none seems to be ancestral to any other subclass, so the earliest monocots may have undergone rapid divergent radiation, establishing five evolutionary lines that have been distinct throughout most of the history of monocots. At present the species of subclass Alismatidae retain the greatest number of relictual features, but no other group appears to have arisen from them. Rather, they have changed least through time, while the other groups have evolved more rapidly and more of their characters have become modified by natural selection.
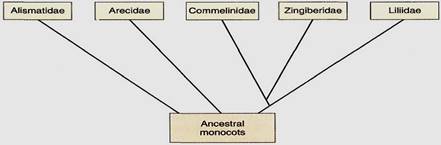
FIGURE 3:Diagram of a widely accepted classification of the monocots, class Liliopsida
Subclass Alismatidae. In the Alismatidae are many aquatic herbs such as Sagittaria (arrowhead; Fig. 4) and many aquarium plants such as Hydrocharis and Naja (Table 1). These plants are most often found in swamps and marshes, partly or entirely submerged. Although many species retain a large number of relictual features, others have become highly modified in response to the unusual selection pressures associated with an aquatic habitat. The plants of many species of the families Hydrocharitaceae, Ruppiaceae, and Najadaceae grow completely submerged in water; they have no transpiration, so mutations that result in the loss of stomata are not selectively disadvantageous. Air chambers make the plants buoyant, so mutations that prevent the formation of lignified fibers are selectively advantageous; the plant does not waste carbon and energy by producing fibers that are not needed for support. Such plants tend to be thin and delicate, all parts of their bodies having very little sclerenchyma and almost no xylem. Tissues have been lost or simplified; the ancestors of Alismatidae were more complex and massive and had more sclerenchyma.
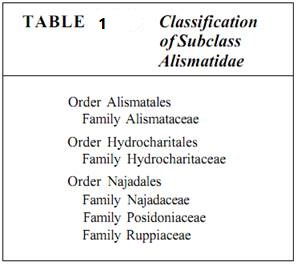

FIGURE 4:Sagittaria, a member of the Alismatidae, is dioecious; plants have either staminate (a) or carpellate (b) flowers.
Other members of subclass Alismatidae have part of their bodies emergent from the water, and some are entirely emergent whenever the marsh dries temporarily. These plants are less highly modified, often having large leaves, considerable amounts of fiber, and a thick cuticle on the leaf epidermis. It is necessary for a species to be adapted to the nest adverse conditions that it frequently encounters during one plant's lifetime. In the most relictual members of Alismatidae, flowers are large and showy, with three sepals and three petals, but in others they arc highly modified, especially in those species in which the flowers do not emerge from the water and pollination occurs below the surface. Sub flowers often completely lack a perianth.
Subclass Arecidae. The Arecidae is a small subclass containing only four orders, five families, and about 5600 species (Table 2). It is made up of larger plants, either aquatic or terrestrial, very often tropical. The most familiar are palms and aroids such as Philodendron, Monstera, and Dieffenbachia.
The family Palmae, also known as Arecaceae, contains about 3500 species, over half the species of the subclass. Palms are easily recognizable by their sturdy solitary trunk and large crown of leaves. In a few species, the trunk is prostrate (palmetto palm) or a vine (Daemonorops). All species have compound leaves, either pinnately compound (feather palms) or palmately compound (fan palms). The typical coconut is one type of palm fruit. Palm flowers are seldom seen because they are usually tiny, about 5 mm across, and are formed only high up in the tree (Fig. 5).
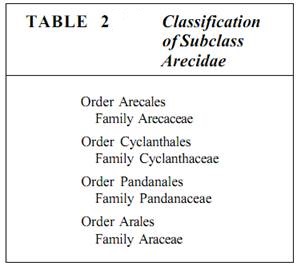

FIGURE 5: (a) Palm flowers usually occur in large inflorescences at the top of a plant. Corypha umbraculifera. (Kjell B. Sandved) (b) Although palm flowers are small, they have typical monocot organization with their parts in sets of three. Syagrus oleracea.
Palms vary somewhat in their body structure, their reproductive biology, and their type of habitat. Consequently they have spread through many tropical and semitropical regions, often being a major aspect of a flora. In most people's minds, palms are synonymous with Florida, Hawaii, and other tropical regions. However, it is instructive to consider palms carefully. The earliest known fossils are 80 million years old, so palms are an ancient group within the monocots. Although in some species the shoot may become massive, they never have any secondary growth. Because the capacity to produce annual, deciduous leaves has not evolved in this family, most palms cannot survive in areas with harsh winters. Because the stems tend to be wide, upward growth is slow, and palms are at a disadvantage when competing for light with trees that can elongate rapidly. As a result, palms are not found in dense forests, but rather in open ones that allow more light to reach the understory plants, such as young palms. Palms are numerous only because they are adapted to a widespread habitat; outside this particular ecological niche, however, they do not thrive.
The Family Araceae (aroids) contains several large orders of familiar houseplants: Philodendron (250 species), Anthurium (500 species), and Arisaema (Jack-in-the-pulpit; 100 species). Dieffenbachia (dumb cane) and 1800 other species also belong here (Fig. 6). This family is characterized by the evolution of a distinctive inflorescence: tiny flowers, either unisexual or perfect, embedded in a thick stalk, the spadix. Staminate flowers are located near the top of the spadix, carpellate flowers near the base. The spadix is surrounded by a large bract, the spathe .

FIGURE 6:This Anthurium (Araceae) is an aroid, one of the monocots that have a dicot type of broad leaf. Many members of this family are adapted to conditions of low light and high humidity; most are members of tropical or subtropical habitats.
Subclass Commelinidae. Subclass Commelinidae contains many familiar groups, such as grasses and sedges (Table 3). The evolution of this group has two prominent aspects. First, the flowers have become adapted to wind pollination (Fig. 7). Only the flowers of the most archaic members are still pollinated by insects (Fig. 8). In almost all other species, pollination is mediated by wind, and structures that would aid in attracting insects, such as nectaries or showy sepals and petals, have been either lost or modified such that they now aid in catching pollen or distributing the fruits after seeds have matured.
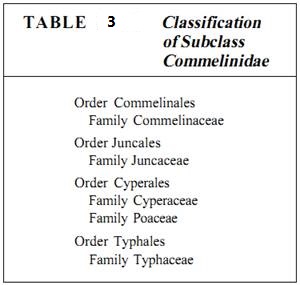

FIGURE 6: This Anthurium (Araceae) is an aroid, one of the monocots that have a dicot type of broad leaf. Many members of this family are adapted to conditions of low light and high humidity; most are members of tropical or subtropical habitats.

FIGURE 7: Grass flowers are very reduced, simplified, and wind pollinated. The usual two small, dry scales (called a lemma and a palea) are thought to be remnants of an ancestral bract and two fused tepals; the third tepal does not form. Three anthers with long filaments and three carpels fused together with long feathery styles and stigmas are usually present. Grass flowers occur grouped together in complex, compact inflorescences, and inflorescence characters are important £r identifying grasses.
The second evolutionary trend is a reduction in body size; most species in the subclass Commelinidae are small herbs. Although they may be fibrous and tough like some grasses and sedges, they are rarely large and massive like palms and philodendrons. The giant bamboos are the only exception.
Members of this subclass fill many ecological niches. They range from tropical species to those that inhabit deserts, grasslands, marshes, forests, and tundra, various species each tolerating extremes of heat, cold, moisture, and drought. Nearly every type of habitat contains at least one member of this subclass.
The spiderwort family, Commelinaceae, contains relatively relictual, showy flowers such as those of Tradescantia, Zebrina, Commelina, and Rhoeo. They have three large sepals and two or three brightly pigmented petals; although they lack nectaries, they are insect pollinated. The androecium has three stamens, and the gynoecium consists of three carpels fused together into a single compound structure.
The grasses, family Poaceae, have about 8000 species and are much more than just the plants in the lawn. They also include most foods, such as wheat (Triticum), barley (Hordeum), oats (Avena), rye (Secale), corn (Zea), rice (Oryza), and sugar cane (Saccharum) as well as a major building material of the tropics, bamboo (the subfamily Bambusoideae). Grasses are extremely abundant in flat, open, dry regions in the central areas of all continents, such as the rangelands of the United States and Africa, the steppes of Russia and Ukraine, and the pampas of Argentina.
All grasses are wind pollinated, so sepals and petals are of little importance and are reduced to bristle-like structures. The three carpels are fused together, but in most grasses the three stigmas remain separate. An "ear" of corn is an inflorescence, and each "silk" is a compound style and stigma . Within the fused ovary of grasses there is just one ovule; once fertilized, it matures into a seed whose seed coat fuses firmly to the developing fruit wall. The objects that we usually call the "seeds" of corn, wheat, and oats are actually single-seeded dry fruits technically known as caryopses (singular: caryopsis). Nearly the entire bulk of the seed is endosperm; the embryo is small but well developed.
Closely related to grasses are sedges (Cyperaceae with 4000 species; Fig. 9) and rushes (Juncaceae). Other members of this subclass are cattails (Typhaceae).

FIGURE 8: Flowers of Commelina have many relictual features. They are still insect pollinated and have a large, showy perianth. There are three petals, but one is smaller than the other two, causing the flower to be bilaterally symmetrical. It often has six stamens, but three may be very small or suppressed. (Robert and Linda Mitchell)

FIGURE 9: Most sedge grow in wet, marshy areas. Carexstricta. (PatLynch/Photo Researchers, Inc.)
Subclass Zingiberidae. This subclass contains some of the most familiar of all house- plants: Maranta, Calathea, canna lilies (Canna), gingers (Zingiber, Hedychium), and bromeliads, as well as some that are best known in the warmer southern states—banana (Musa) and bird-of-paradise (Strelitzia) (Table 4). Members of subclass Zingiberidae differ from those of the other monocot subclasses discussed so far in that they tend to have large, showy flowers pollinated by insects, birds, or bats. Furthermore, many of flowers have derived features; adjacent sepals are often fused to each other, forming a tube, and the same is often true of the petals. The gynoecium typically consists of three carpels that have fused almost completely, except for small regions that act as nectaries (septal nectaries). Very often the gynoecium is inferior, located below the sepals and petals, a condition that is always considered quite derived, the result of considerable evolutionary modification.
Subclass Zingiberidae contains two orders (Bromeliales and Zingiberales), nine families, and almost 3800 species. Members of one subclass of monocots often resemble those of other subclasses; the Zingiberidae show that it can be quite difficult to be certain which groups are related to which. The Bromeliales are often classified with the Commelinidae, and the Zingiberales with the Liliidae. Not everyone would agree that the two orders should be combined and placed in their own subclass, as has been done by Dr. Cronquist. Many characters are being analyzed to understand the phylogeny of these groups. In addition to morphology and anatomy, important features are the nature of crystals, starch grains, silica deposits, protein bodies, chemical characters, and even susceptibility to attack by certain fungi. It is difficult to know which features are homologous (descent from a common ancestor) and which are analogous (convergent evolution). Until definitive proof of one classification or another can be obtained, we must be aware of the complexity and understand why differences of opinion exist.

Order Bromeliales has only one family, Bromeliaceae, which contains some of the most beautiful tropical epiphytes, their large, brightly colored inflorescences being easily visible even in thick jungle vegetation (Fig. 25.19a). Epiphytic species extend as far northward as the subtropics, Spanish moss and ball moss (both are species of Tillandsia) occurring from Florida to Texas (Fig. 10b). Other species are terrestrial and usually xerophytic, such as Puya of Chile and Peru and pineapples (Ananas). These xerophytic terrestrial forms are believed to be relictual, and their capacity to withstand drought may have made it easier for bromeliads to adapt to life as epiphytes. Because bromeliads occur only in the Americas, it is assumed that they evolved after South America separated from Africa about 80 million years ago . Had they evolved earlier, numerous species should occur in the African rain forests and coastal deserts.
Order Zingiberales contains eight families; the largest, Zingiberaceae, has about 1000 species. Members of this order are almost all tropical, and most are soft, nonleathery herbs. The leaves are broad and have a petiole, resembling those of dicots, but they are not believed to be homologous with dicot leaves (Figs 11). During the evolution of the Zingiberales, mutations that resulted in the petiole/leaf becoming flattened and broadened were selectively advantageous: The plants grow in the heavy shade of jungle understory where light interception is important.

FIGURE 10: (a) The inflorescences of many bromeliads have brilliant pigmentation. One hypothesis postulates that this coloration increases visibility of the flowers in the dim light of a dense rain forest. (D. Cavagnaro/Visuals Unlimited) (b) Spanish moss, Tillandsia usneoides, is a bromeliad, not a moss. It grows attached to tree trunks as an epiphyte, not a parasite. This species grows in the southeastern United States, where humidity is high and rainfall is frequent. (c) Pineapple plants are terrestrial bromeliads; they grow with their roots in soil. The edible portion of pineapple is a multiple fruit, composed of an entire inflorescence. (Phil Degginger).

FIGURE 11: Some monocots, such as this banana (Musa), have broad leaves, but their lamina is not homologous to that of a dicot leaf. In this monocot broad leaf, all vascular bundles run parallel to each other from the midrib to the margin, whereas in a dicot lamina, they diverge and branch into a reticulate pattern.
Subclass Liliidae. Most botanists consider subclass Liliidae to contain the most derived of all monocots, lilies (family Liliaceae with 4000 species) and orchids (Orchidaceae with over 20,000 species) plus 17 other families (Table 25.7). Most species of Liliidae rely on insect pollination and have large, showy flowers that are often wonderfully aromatic (Figs. 12 and 13a). Septal or other types of nectaries are usually well developed. A common habit (growth form) is the formation of a tough succulent stem; in the lily family the stem is often subterranean as a corm, rhizome, or bulb. In orchids it is frequently present as an aerial pseudobulb that can persist even during stressful conditions. The leaves and even roots may die, but when conditions become favorable, the pseudobulb produces new leaves, roots, and flowers.
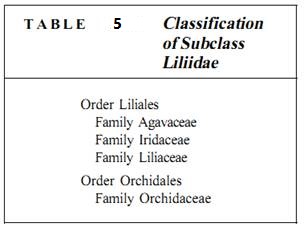

FIGURE 12: Lilies and their relatives are excellent for demonstrating the typical monocot flower: They have three of each type of appendage and all are large enough to be seen easily. The three carpels are fused together, but a transverse section shows three chambers. (Runk/Schoenberger from Grant Heilman)

FIGURE 13: Members of subclass Liliidae. (a) Iris. (b) Like bamboos and palms, yuccas and agaves prove that herbs do not have to be soft and small. The leaves are long and narrowly triangular with parallel venation, and they may be thick at the base. They are arranged as a rosette around a short (agaves) or long (yuccas) stem. Most have a needle-sharp point and razor-like hooked spines along the edges of the leaf. (Courtesy of David Bogler, University of Texas)
plants that most people are familiar with them and think of them as the "typical" monocots. Liliaceae contains lilies, day-lilies, tulips, onions, asparagus, and amaryllis. The heavier, coarser agaves and yuccas are sometimes placed in the Liliaceae, sometimes in their own family, the Agavaceae. The lily family is believed to have given rise to the Iridaceae (irises) and Dioscoreaceae (yams).
The Orchidaceae is the largest and most diverse family of all monocots. The most familiar, ornamental ones are epiphytic, but many are terrestrial and one is a subterranean parasite. Orchid flowers are highly modified from the ancestral condition of the angiosperms, being zygomorphic (bilaterally symmetrical) with complex shapes, colors, and fragrances that attract specific pollinators. Orchids are unusual in producing hundreds or thousands of tiny seeds in each fruit. The seeds are dust-like and so undeveloped at "germination" that they must form a symbiosis with certain fungi in order to survive long enough to form roots and leaves.
According to Dr. Cronquist, the orchids must be derived from lilies; the most derived lilies have characters that would be expected to occur in the ancestors of orchids. Virtually all features of the orchids arc highly derived and extremely modified. They have little in common with those members of subclass Alismatidae that are thought to retain relictual features of the earliest monocots.



|
|
|
|
التوتر والسرطان.. علماء يحذرون من "صلة خطيرة"
|
|
|
|
|
|
|
مرآة السيارة: مدى دقة عكسها للصورة الصحيحة
|
|
|
|
|
|
|
نحو شراكة وطنية متكاملة.. الأمين العام للعتبة الحسينية يبحث مع وكيل وزارة الخارجية آفاق التعاون المؤسسي
|
|
|
















