
آخر المواضيع المضافة

 النبات
الحيوان
الأحياء المجهرية
علم الأمراض
التقانة الإحيائية
التقنية الحيوية المكروبية
التقنية الحياتية النانوية
علم الأجنة
الأحياء الجزيئي
علم وظائف الأعضاء
الغدد
المضادات الحيوية
النبات
الحيوان
الأحياء المجهرية
علم الأمراض
التقانة الإحيائية
التقنية الحيوية المكروبية
التقنية الحياتية النانوية
علم الأجنة
الأحياء الجزيئي
علم وظائف الأعضاء
الغدد
المضادات الحيوية| الأليلات المتعددة Multiple Alleles |


|
|
أقرأ أيضاً
التاريخ: 13-11-2015
التاريخ: 12-11-2015
التاريخ: 13-11-2015
التاريخ: 12-11-2015
|
الأليلات المتعددة Multiple Alleles
بديهي إن أقصى عدد للأليلات لموقع جيني في الكائن هو إثنان ، واحد على كل من الكروموسومات المتماثلة ((Homologous chromosomes وقد اقتصرت الأمثلة الوراثية السابقة على التعامل بزوج واحد من الأليلات للجين الواحد مثل AA و Aa و aa . وبما إن تغيير الجين إلى أشكال بديلة أخرى ((Alternative forms يتم عن طريق الطفرات فمن الممكن نظرياً حدوث طفرات متعددة في الجين ، وتسمى هذه الطفرات المتعددة في الجين الواحد بالأليلات المتعددة إذا ما سببت تغييراً في النمط الظاهري البري للكائن .
الأليلات المتعددة في الدروسوفلا :
يتحكم بلون عين الدروسوفلا عدد من الأليلات التي تسبب تبايناً في اللون من الأحمر في النوع البري وشفرته الأليل (w+) إلى اللون القرنفلي (w co) والأحمر القاني (w bl) والوردي (w e) والأحمر الفاتح (w ch) والمشمشي (w a) والعسلي (w h) والأصفر البرتقالي (w bf) والبرتقالي الفاتح (w t) واللؤلؤي (w p) والعاجي (w i) والأبيض (w) . إن هذه الأليلات قادرة على أن تنتج صبغة ما عدا الأليل (w) ويمكن توضيح كمية الصبغة المنتجة من قبل هذه الأليلات بالصورة التالية :-
w+ ˃ w co ˃ w al ˃ w e ˃ w ch ˃ w a ˃ w h ˃ w bf ˃ w t ˃ w p ˃ w i ˃ w
حيث إن الأليل البري (w+) يكون ذو سيادة كاملة على كل الأليلات الأخرى في هذه السلسلة ، أما الأليل (w) فيكون متنحياً لكل الأليلات الأخرى في هذه السلسلة الأليلية ، وعندما تكون أليلات هذه السلسلة ( ما عدا w+ ) متباينة الزيجة (Heterozygote) فإنها تميل إلى إعطاء نمط ظاهري متوسط (Intermediate) بين ألوان عين الدروسوفلا الأبوية .
وكما لاحظنا في المثال السابق فإن الأليلات المتعددة تسبب تدرجاً في اللون لنمط ظاهري معين ، ولكن هناك أليلات لجينات أخرى تنتج أنماطاً ظاهرية مختلفة .
فالجين (ss) في الدروسوفلا ميلانوجستر يسبب الاهلاب الصغيرة (Small bristles) على عكس النمط الظاهري البري للجين (ss+) . ولم يلاحظ أي تأثير من قبل الجين (ss) على الأرجل (Legs) ولا على قرون الإستشعار (Antennae) . وهناك أليل آخر في هذه السلسلة الأليلية وهو الأليل (ssa) (Aristapedia) الذي يختزل الأهلاب قليلاً إلا إنه يسبب تغييرات أكثر وضوحاً في النمط الظاهري حيث تنشأ أرجلاً على الرأس بدلاً من قرون الإستشعار .
وراثة لون فراء الأرنب :
يتحكم في لون الفراء البري للأرنب الجين (C) والذي يعطي اللون الرمادي ويسبب الأليل (cch) لون الشنشلا (Chinchila) عندما يكون متماثل الزيجة (Homozygous) وأما في حالة كونه متباين الزيجة (Heterozygous) مع أليل ذو درجة أقل في سلم السيادة فإنه ينتج اللون الرمادي الفاتح ، أما الأليل (ch) فينتج الأرنب الأبيض ذو النهايات السوداء وتسمى هذه الحالة الهيمالايان (Himalayan) ، أما الأليل (c) فإنه لا يستطيع إنتاج أي صبغة مؤدياً الى الأرنب الأمهق (Albino) , ويعتبر هذا الأليل متنحياً لكل الأليلات ونستطيع أن نمثل السيادة بين هذه الأليلات كما يلي :
C< cch < ch < c
وجدول رقم (1) يبين الأنماط الظاهرية العائدة للأنماط الوراثية المختلفة .
جدول رقم (1) الأنماط الوراثية المختلفة المسببة للأنماط الظاهرية للون فراء الأرنب .
|
النمط الظاهري |
النمط الوراثي |
|
الفراء البري الرمادي |
CC , Ccch , Cch , Cc |
|
الشنشلا |
cchcch |
|
الرمادي الفاتح |
cchch , cchc |
|
الهيمالايان |
chch , chc |
|
الأمهق |
cc |
مجاميع الدم ABO في الإنسان :
في مطلع هذا القرن لاحظ لاندشتاينر (K. landsteiner) حصول تجمع لكريات الدم الحمراء (Red corpuscles) لشخص عند خلطها مع مصل الدم الذي يعود لشخص آخر , وبعد عمل عدة تجارب تشمل عينات دم من أفراد عديدين إستطاع هذا العالم أن يصنف الدم في الإنسان الى ثلاثة مجاميع وهي A و B و O .
ومن ثم أكتشف فون دي كاستيلو Von Decastello)) وستارلي Sturli)) وهما من تلاميذ لاندشتاينر مجموعة الدم (AB) . وهذا التقسيم لمجاميع الدم في الإنسان يعود الى إن بعض الأفراد يملكون أما المستضد (Antigen) A أو المستضد (B) وآخرون يملكون كل من المستضدين (A و B) معاً وآخرون لا يملكون أياً منهما , إن الأفراد من المجموعة (A) لا يحملون في المصل الضد A- (Anti-A) ولكن يحملون الضد B- (Anti-B) أما الأفراد من مجموعة الدم (B) فيحملون الضد A- ولا يحملون الضد B- , لهذا السبب يتوقع حدوث تجمع لكريات الدم الحمراء عند نقل الدم من الفرد A الى الفرد B أو من B الى A . ومثل هذا التفاعل يحصل عند خلط الدم من المجموعة A أو B مع الأمصال المضادة (Antisera) في أنبوبة الأختبار ، لذلك يمكن إجراء الفحوص اللازمة للدم مختبرياً قبل عملية نقل الدم , والأفراد للمجموعة AB يملكون المستضدات A و B في كريات الدم الحمراء ولكن لا يملكون الضدين A و B في المصل , وأما الأفراد من المجموعة O فلا يملكون المستضدات A و B ولكن يحملون الضدين A و B . ويمكن نقل الدم من أفراد المجموعة O الى أي فرد في المجاميع A و B و AB بشرط أن يتم نقل الدم ببطء مناسب ليسمح بالتخفيف السريع للضدين A و B في مصل المعطي (Donor) وذلك في دم الآخذ (Recipient) .
إن السلوك الوراثي للجينات المسؤولة عن مجاميع الدم A و B و AB و O وضع على أساس سلسلة من ثلاثة أليلات لجين واحد وهو I . حيث يرمز الأليل IA للمستضد A ويعتبر متساوي السيادة (Codominant) مع الأليل IB الذي يمثل المستضد B ويعتبر كل من الأليلين IA و IB ذو سيادة كاملة على أليل (i) الذي لا يعين أي مستضد , ويمكن أن تمثل علاقة السيادة بين هذه الأليلات كالآتي :
IA = IB <i
والجدول رقم (2) يوضح الأنماط الوراثية المختلفة لمجاميع الدم .
جدول (2) الأنماط الوراثية المختلفة لمجاميع الدم A و B و AB و O .
|
النمط الوراثي |
النمط المظهري |
|
IAi , IA IA |
A |
|
IBi , IB IB |
B |
|
IA IB |
AB |
|
ii |
O |
مجاميع الدم ريسس (Rhesus) في الإنسان:
بعد إنتشار عملية نقل الدم على نطاق واسع كنتيجة لإكتشاف مجاميع الدم ABO حصلت عدد من حالات تجمع كريات الدم بسبب نقله من فرد الى آخر بالرغم من تطابق مجموعة الدم ABO . وأكتشف في عام 1940م العامل Rh من قبل كل من العالمين لاندشتاينر وفاينر Landsteines and Wiener) ) وذلك في دم أرانب سبق وإن نقل اليها دم قردة Macaca rhesus وأدت الأجسام المضادة المتكونة في دم الأرانب الى تجمع كريات الدم الحمراء ليس في القردة فقط ولكن في نسبة محسوسة من المجموعة البشرية القوقازية ايضاً . وعرف الأفراد الحاملين لهذا العامل في كريات الدم الحمراء بـ Rh+ أما غير الحاملين لهذا العمل فرمز له Rh- . ويختلف المستضد Rh عن مستضدات مجموعة الدم AB بعدم وجود الأجسام المضادة في مصل الافراد من نوع Rh- . ولكن بإمكان مثل هؤلاء الأشخاص (Rh-) من تكوين الأجسام المضادة إذا ما تم نقل الدم اليهم من أفراد (Rh+) . وستؤدي عملية نقل الدم ما بعد الأولى من شخص (Rh+) الى آخر (Rh-) سبق وتكونت في دمه الأجسام المضادة الى تجمع كريات الدم الجمراء التي تعود للواهب وبالتالي موت الفرد المستلم . لذلك يتم في الوقت الحاضر التأكد من نوع العامل Rh إضافة الى نوع مجاميع الدم ABO قبل إجراء عملية نقل الدم . وكنتيجة لإكتشاف عامل ريسس إنتهت إحدى المشاكل الطبية المستعصية . فكما هو معروف يولد سنوياً كثير من الأطفال مصابين بمرض (Erythroblastosis Fetalis) ويسمى هذا المرض أيضاً مرض فقر الدم الأنحلالي Hemolytic disease وقسم من الأفراد المصابين يجابهون المنية قبل الولادة . وبينت التحاليل إن العامل Rh يكون في الآباء والأطفال موجباً وفي الأمهات سالباً . وهذا يشير الى إن النمط المظهري الموجب للعامل Rh هو السائد .
ولا يحدث هذا المرض في الطفل الأول . ولكن عند فصل المشيمة عن جدار الرحم خلال عملية الولادة الأولى يحدث نزيف بكمية تسمح بدخول عدد من كريات الدم الحمراء من الوليد والحاملة للمستضد من نوع Rh+ الى دم الأم . وهذا يؤدي الى تكوين الأجسام المضادة نتيجة وجود المستضدات الغريبة .
وتضاربت الدراسات حول الأسس الوراثية للعامل Rh . فقد أقترح فاينر (Wiener) وجود ثمانية أليلات في الموقع الجيني Rh وأزداد عدد الأليلات في هذا الموقع الجيني منذ أكتشافها . أما فيشر (Fisher) فقد أقترح نظرية وراثية أخرى تستند على نظرية الأليلات الكاذبة (Pseudoalleles) وبين وجود ثلاث مواقع جينية تقع قريبة من بعضها على الكروموسوم ومسؤولة عن ثلاث مستضدات مختلفة ويرمز لهذه المستضدات في الأفراد من نوع Rh+ بالحروف C و D و E وأما الرموز c و d و e فتشير الى الأفراد Rh- . والفرد الحامل لأحدى الجينات C أو D أو E أو إثنان أو بمجموعها يكون من فئة Rh+ أما الفرد من فئة Rh- فيكون حاملاً للجينات c و d و e أي إن الفرد يجب أن يكون في حالة متماثل الزيجة لكل الأليلات المتنحية . وعند فحص مجاميع الدم Rh يستعمل المضاد المصلي (Antiserum) من نوع Anti-CDE فإذا ما كان الفرد حاملاً لواحد أو أكثر من المستضدات فإن كريات الدم ستتجمع ويعتبر D Rh أكثر المستضدات شيوعاً حيث تصل نسبة الأفراد Rh+ الحاملين بهذا النوع من المستضدات الى 98٪ , وتبين أيضاً إن هذا المستضد هو أكثر مستضدات مجموعة Rh فعالية .
أليلات العقم الذاتي في النباتات Self Sterility Alleles
توجد الأليلات المتعددة لظاهرة العقم الذاتي في عدد من النباتات مثل الاينوثيرا المغربية (Evening primroses) والتبغ (Tobacco) والبرسيم (Clovre) والكرز (Cherries) وتعرف هذه الأليلات بالسلسلة S1 ، S2 ، S3 . . . . إلخ . ومثال ذلك حبة لقاح التبغ الأحادية المجموعة الكروموسومية الحاملة لأليل العقم الذاتي (S1) التي لا تستطيع النمو جيداً على القلم الثنائي المجموعة الكروموسومية (Diploid female style) ذا النمط الوراثي S1S2 حيث إنها حاملة لنفس الأليل S1 ، ولكن تستطيع حبة اللقاح هذه النمو بنجاح على القلم الثنائي للمجموعة الكروموسومية ذا النمط الوراثي S2 S3 أو S3 S4 . . . إلخ . ومن هذا نستنتج إن الإخصاب الناجح (Successful fertilization) في عدد من الحالات في النبات أعلاه إنما يدل على وجود أليل في حبة اللقاح الذي يختلف عن كل من الأليلين الحاضرين في القلم الثنائي للمجموعة الكروموسومية .
التركيب الدقيق للجين :
إن ملاحظة وجود أليلات متعددة للجين قد مهدت الطريق لدراسة مواقع هذه الأليلات في داخل الجين وبالتالي إمكانية رسم الخارطة الجينية ، ولأجل دراسة التركيب الدقيق للجين يجب توفر سلالات (طفرات) مختلفة وعديدة تقع في نفس الجين ويتم جمع هذه الطفرات وفق الشروط التالية :
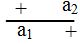 ولكن يحدث التكميل (Complementation) في سيس متباينة الزيجة (Cis Heterozygote)
ولكن يحدث التكميل (Complementation) في سيس متباينة الزيجة (Cis Heterozygote)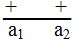 .
.
إن هذه الشروط يمكن ملاحظتها وتطبيقها على عدد من الطفرات في الكائنات حقيقية النواة .
التركيب الدقيق للجين ad في العفن Asperigilus Nidulans
لدراسة التركيب الدقيق لإحدى الجينات في (A. Nidulans) تم عزل طفرات ذات عوز غذائي للأدنين (أي إن هذه الطفرات لا تستطيع تخليق الأدنين) وهذه الطفرات هي 8 ad ، 10 ad ، 11 ad ، 16 ad ، وهذا يعني توفر الشرط الأول لدراسة التركيب الدقيق للجين ، ويتوفر الشرط التالي أيضاً في هذه الطفرات من خلال التحليلات الوراثية حيث أوضحت إن هذه الطفرات تقع على نفس الذراع الكروموسومي وما بين العلامة الوراثية (Genetic marker) y (شفرة الكونيديا الصفراء) والعلامة bi (شفرة العوز الغذائي للبايوتين) , وكل هذه الطفرات تكون أقرب إلى العلامة y . ويتوفر الشرط الثالث في هذه الطفرات فعند تضريب الطفرة ad8 مع الطفرة ad16 ينتج نسل نادر نمطه الظاهري بري غير محتاج للأدنين ويستطيع النمو على الوسط الغذائي الأدنى (وعلى هذا الوسط لا يمكن ملاحظة الناتج المتبادل (Reciprocal product) إلا وهو (ad8 ad 16) ويعمل كل التضريبات الممكنة بين الطفرات الأربعة (Pairwise combination) وحساب تكرار التراكيب الجديدة الناتجة . وإن ترتيب الأليلات هو الآتي :
ad 16 ad 11 ad 8 ad 10 di
إن هذه النتائج قد تشير الى وجود جين واحد (ad) ذو أليلات متعددة . أو أربعة جينات مرتبطة أرتباطاً وثيقاً مع بعضها ومسؤولة عن صنع الأدنين , ومن الممكن ترجيح كفة أحد الخيارين أعلاه بعمل إختبار التكميل (Complementation) فقد وجد إن الهيتروكاريون (Heterokaryon) الناتجة من السلالتين ad 8 و ad 16 تبقى ذات عوز غذائي للأدنين , وعندما تعزل الزيجة الثنائية للمجموعة الكروموسومية ذات النمط الوراثي 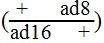 من الهيتروكاريون فإنها أيضاً تبقى بحاجة للأدنين , وفقط عندما يكون النمط الوراثي للزيجة الثنائية للمجموعة الكروموسومية في حالة سيس (Cis)
من الهيتروكاريون فإنها أيضاً تبقى بحاجة للأدنين , وفقط عندما يكون النمط الوراثي للزيجة الثنائية للمجموعة الكروموسومية في حالة سيس (Cis)  فإنها سوف لن تحتاج الى الأدنين .
فإنها سوف لن تحتاج الى الأدنين .
من هذا نرى بأن الطفرات ad 8 , ad 10 , ad 11 و ad 16 هي طفرات أليلية (Allelic mutation) والتراكيب الجديدة (Recombinants) الحاصلة من تضريب الطفرات فيما بينها تأتي عن طريق التركيب الجديد للأليلات البينية (Intrallelic recombination) وطبيعي فإن مجموعة الأليلات الموجودة في هذا الموقع (الجين) تسمى الأليلات المتعددة (Multiple alleles) . وعند رسم الخريطة لجين بطريقة التركيب الجديد (Recombination) يلاحظ إن عدداً من الأليلات تقع في موقع واحد وتسمى الأليلات المتماثلة (Homoalleles) وعند تواجد الأليلات في مواقع مختلفة من الجين تسمى الأليلات المتباينة (Heteroalleles) والأليلات في هاتين المجموعتين تكون متشابهة في الحاجة للأدنين كما في المثال أعلاه , ولكن يمكن تمييزها عن بعضها بخواصها الطفرية وخواصها في إختبار التكميل وحساسيتها للحرارة وللـ pH , ولتوضيح ذلك نأخذ مثال وهو الإختلاف في الأنماط الظاهرية للطفرات الأليلية في موقع ad 6 لخميرة سكيزومايسيس بومبي (Schizosaccharomyces pombe) حيث إن كل طفرات العوز الغذائي للأدنين المختلفة لهذه الخميرة تقع في جين واحد ad 6 ولغرض توضيح الإختلاف في طبيعة هذه الطفرات فقد زود الوسط الغذائي الأدنى في الطبق البتري الأول بـ 75 مايكروغرام /مل من الأدنين وهو تركيز كافي لتثبيط التغذية الإسترجاعية (Feedback inhibition) ولكي تنمو الطفرات نمواً طبيعياً ويكون بالتالي نمطها الظاهري برياً أما في الطبق البتري الثاني فيحتوي على 10 مايكروغرام/مل من الأدنين وهذا التركيز يسهل التمييز مظهرياً بين الطفرات المختلفة في هذا الجين , حيث تستطيع الطفرات النمو ولكن لا يستطيع الأدنين بهذا التركيز من تثبيط التغذية الإسترجاعية بصورة كاملة (Complete feedback inhibition) مما يسبب ظهور فعالية الناتج الجيني لـ ad 6 للأليلات المختلفة معطياً أنماطاً ظاهرية مختلفة تبعاً لفعالية البروتين المتأثر بالطفرة أما على الوسط الغذائي الأدنى فلا تستطيع هذه الطفرات من النمو أي أن لها نمطاً ظاهرياً مماثلاُ .



|
|
|
|
تفوقت في الاختبار على الجميع.. فاكهة "خارقة" في عالم التغذية
|
|
|
|
|
|
|
أمين عام أوبك: النفط الخام والغاز الطبيعي "هبة من الله"
|
|
|
|
|
|
|
المجمع العلمي ينظّم ندوة حوارية حول مفهوم العولمة الرقمية في بابل
|
|
|
















