
آخر المواضيع المضافة

 النبات
الحيوان
الأحياء المجهرية
علم الأمراض
التقانة الإحيائية
التقنية الحيوية المكروبية
التقنية الحياتية النانوية
علم الأجنة
الأحياء الجزيئي
علم وظائف الأعضاء
الغدد
المضادات الحيوية
النبات
الحيوان
الأحياء المجهرية
علم الأمراض
التقانة الإحيائية
التقنية الحيوية المكروبية
التقنية الحياتية النانوية
علم الأجنة
الأحياء الجزيئي
علم وظائف الأعضاء
الغدد
المضادات الحيوية| Igs Expressed in Avians Are Assembled from Pseudogenes |


|
|
Read More
Date: 10-5-2016
Date: 3-12-2015
Date: 4-5-2016
|
Igs Expressed in Avians Are Assembled from Pseudogenes
KEY CONCEPTS
- An Ig gene in chickens is generated by copying a sequence from one of 25 pseudogenes into the recombined (acceptor) V gene (i.e., gene conversion).
- The enzymatic machinery of gene conversion depends on activation-induced deaminase (AID) and enzymes involved in homologous recombination.
- Ablation of certain DNA homologous recombination genes transforms gene conversion into somatic hypermutation (SHM).
The chicken Ig locus is the paradigm for the Ig somatic diversification mechanism utilized by rabbits, cows, and pigs; that is, gene conversion. A similar mechanism is used by both the single (λ-like) L chain locus and the H chain loci. The chicken λ locus comprises only one functional Vλ gene segment, one Jλ segment, and one Cλ gene segment (FIGURE 1). Upstream of the functional Vλ 1 gene segment lie 25 Vλ pseudogenes, organized in either orientation. In the pseudogenes, either the coding segment is deleted at one or both ends or proper RSSs are missing, or both.
This is emphasized by the fact that only the Vλ 1 gene segment recombines with the Jλ -Cλ gene segment.
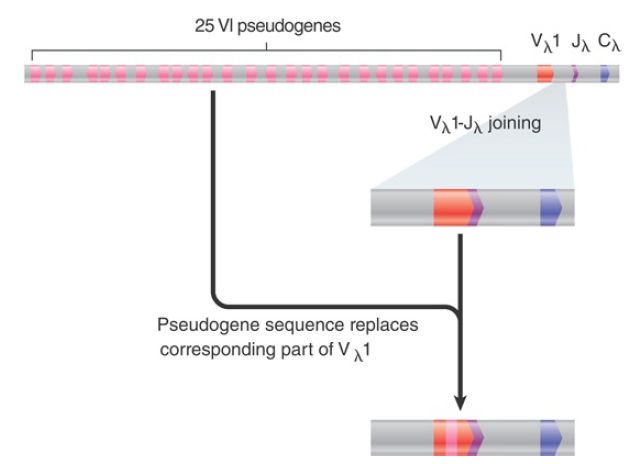
FIGURE 1 The chicken λ light chain locus has 25 V pseudogenes upstream of the single functional Vλ -Jλ -C region. Sequences derived from the pseudogenes, however, are found in active rearranged VJC genes.
Nevertheless, sequences of rearranged Vλ Jλ -Cλ gene segments show considerable diversity. A rearranged gene has one or more positions at which a cluster of changes occurred in its sequence. A sequence identical to the new sequence can almost always be found in one of the pseudogenes. The sequences that are not found in a pseudogene always represent changes at the junction between the original sequence and the altered sequence. The unmodified Vλ 1 sequence is not expressed, even at early times during the immune response. Sequences from the pseudogenes, between 10 and 120 bp in length, are integrated into the active Vλ 1 region by gene conversion. A successful conversion event probably occurs every 10 to 20 cell divisions to every rearranged Vλ 1 sequence. At the end of the immune maturation period, a rearranged Vλ 1 sequence has four to six converted segments spanning its entire length, which are derived from different donor pseudogenes. If all pseudogenes can participate in this gene conversion process, more than 2.5 × 108 possible combinations are allowed.
The enzymatic basis for copying pseudogene sequences into the recombined Ig V gene depends on AID and enzymes involved in homologous recombination, and is related to the mechanism of human and mouse SHM . For example, gene conversion is prevented by deletion of RAD54. Deletion of other homologous recombination genes, such as XRCC2, XRCC3, and RAD51B, has another interesting effect: Somatic mutations occur in the V gene of the expressed locus. The frequency of the somatic mutations is 10-fold greater than the rate of gene conversion.
Thus, the absence of SHM in chicken is not due to a deficiency in the enzymatic machinery that is responsible for SHM in humans and mice. The most likely explanation for a connection between (lack of) recombination and SHM is that unrepaired DSBs in the recombined Ig V(D)J segments trigger the induction of mutations. The reason why SHM occurs in mice and humans but not in chickens may, therefore, lie with the nature of the repair system that operates on DSBs in the Ig locus. It would be more efficient in chickens, so that DSBs in the Ig locus are repaired through gene conversion before mutations can be induced.



|
|
|
|
تفوقت في الاختبار على الجميع.. فاكهة "خارقة" في عالم التغذية
|
|
|
|
|
|
|
أمين عام أوبك: النفط الخام والغاز الطبيعي "هبة من الله"
|
|
|
|
|
|
|
قسم شؤون المعارف ينظم دورة عن آليات عمل الفهارس الفنية للموسوعات والكتب لملاكاته
|
|
|
















