
آخر المواضيع المضافة

 النبات
الحيوان
الأحياء المجهرية
علم الأمراض
التقانة الإحيائية
التقنية الحيوية المكروبية
التقنية الحياتية النانوية
علم الأجنة
الأحياء الجزيئي
علم وظائف الأعضاء
الغدد
المضادات الحيوية
النبات
الحيوان
الأحياء المجهرية
علم الأمراض
التقانة الإحيائية
التقنية الحيوية المكروبية
التقنية الحياتية النانوية
علم الأجنة
الأحياء الجزيئي
علم وظائف الأعضاء
الغدد
المضادات الحيوية| Membranes |


|
|
Read More
Date: 8-11-2016
Date: 18-10-2016
Date: 29-10-2016
|
Membranes
All cells contain at least some membranes, and cells of eukaryotes (plants, animals, fungi, and protista) contain numerous organelles composed of membranes. Membranes perform many important tasks in cell metabolism: They regulate the passage of molecules into and out of cells and organelles; they divide the cell into numerous compartments, each with its own specialized metabolism; and they act as surfaces that hold enzymes. Without membranes, life would be impossible; indeed, many things that cause death in both humans and plants—heat, cold, many poisons, alcohol—are able to kill because they disrupt membranes.
COMPOSITION OF MEMBRANES
All biological membranes are composed of proteins and two layers of phospholipid molecules (see 1). If phospholipids are poured carefully onto the surface of calm water, they automatically form a single layer (a monolayer) on the water's surface, with their hydrophilic ends forming hydrogen bonds to water molecules and their hydro- phobic fatty acids projecting out of the water (Fig. 2). If the water is agitated, the lipid layer doubles over and makes a bilayer in which all fatty acids are away from the water and all phosphate groups are in full contact with it. Any break or tear in a bilayer membrane exposes hydrophobic fatty acids to water, so membranes always reseal themselves after a rupture.

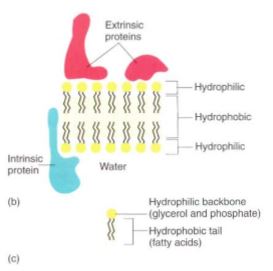
FIGURE 1: (a) In electron micrographs, membranes appear as two dark lines (proteins) separated by a light region (composed mostly of lipids) (X 600,000). (b) Pictures such as (a) led scientists to believe that membranes consist of two layers of lipids with proteins located only on the surface. However, many proteins fit partly into the lipid bilayer or pass all the way through it. (c) A phospholipid has three parts: (1) two fatty acids, (2) a backbone of glycerol, and (3) a phosphate group.
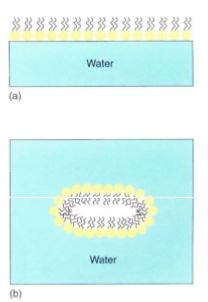
FIGURE 2: (a) In quiet water, phospholipids form a monolayer at the surface; by projecting away from the water, the fatty acids do not disrupt the hydrogen bonding between water molecules. (b) With agitation phospholipids form a bilayer; this arrangement allows the greatest number of hydrogen-bonds to form between phosphates and water while simultaneously minimizing the disruption of water-water hydrogen-bonds by the fatty acids.
The lipid bilayer is a very thin solution. If it contains several types of lipid, they can diffuse laterally throughout the membrane because they are not bonded to each other, but they cannot diffuse vertically from the membrane into the surrounding solution. All biological membranes contain proteins as well as lipids, usually in a ratio of 60% proteins and 40% lipid (see Fig. 1b). Most proteins have large hydrophilic regions, so they associate mostly with the phospholipid phosphates and with water. Many also have large hydrophobic regions that allow them to sink into the membrane and associate with the fatty acids (Fig. 3). Variations in hydrophobic and hydrophilic regions mean that various proteins sit entirely on the membrane surface, are partly immersed in it, or span it entirely, with either end projecting out of opposite sides. This allows the protein to have its active site on either or both sides of the membrane or within it. Likewise, a protein may act as a hydrophilic channel that permits small hydrophilic molecules to pass through the membrane. Proteins that are even partially immersed in the lipid bilayer are said to be intrinsic proteins. Others, extrinsic proteins (also called peripheral proteins), are located outside the membrane and merely lie next to it. Although extrinsic proteins may perform important enzymatic functions, they are not an integral part of the membrane's structure. Some intrinsic proteins contribute to the membrane's fluid nature and, like the lipids, can diffuse laterally. Other proteins interact with adjacent proteins, forming complexes or domains (small discrete regions) different from surrounding regions of the membrane. If all membrane components could freely diffuse laterally, the membrane would become homogeneous and no differentiation could occur. But because at least some proteins are bound to their neighbors, membranes are heterogeneous and patchy, and differentiation does take place. Because the membrane is a heterogeneous liquid, it is said to be a fluid mosaic.
Some membranes contain a small amount of sugar, usually less than 8%. The sugars occur as short-chain oligosaccharides, each with about 4 to 15 sugar residues. These oligosaccharides are bound to certain intrinsic proteins, converting them into glycoproteins; rarely sugars are attached to membrane lipids (glycolipids). At present, we think that these polysaccharides make the membrane more distinctive and easy to recognize, an especially important feature in animals that have an immune system. Specialized protective cells attack and destroy anything they do not recognize as a part of the animal's own body. The glycoproteins and glycolipids occur almost exclusively on the outer surface of the membrane that covers the cell, a position that allows maximum exposure for these polysaccharides. Glycoproteins and glycolipids may be less important in plants.
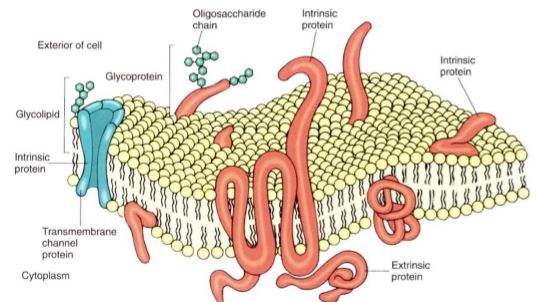
FIGURE 3: Intrinsic membrane proteins differ in their size, shape, and location of hydrophobic, lipid-soluble regions; consequently, some sink deep into the membrane, others span it, and some penetrate only one lipid layer.
PROPERTIES OF MEMRRANES
Membranes have several important properties. First, they can grow. Membranes are formed molecule by molecule in certain regions of the cell; then whole pieces of membrane are moved as small bubbles or vesicles to different sites in the cell. When the vesicle of preformed membrane arrives at the growing membrane, the two fuse (Fig. 4).
In addition to permitting movement of membrane pieces, membrane fusion allows the transport of material. The volume inside the vesicle (the vesicle lumen) may be filled with substances that must be accumulated, broken down, or otherwise metabolized at the vesicle's destination. Vesicle movement may also release material to the outside of the cell (Fig. 4). This exocytosis may be a means to excrete almost anything: wastes, debris, mucilage, proteins, polysaccharides. For example, roots slide through the soil by secreting a slippery, lubricating mucilage that is formed within the root cells, packaged into vesicles that migrate to and fuse with the cell surface membrane, and are then released to the exterior. In many flowers, nectar is secreted from glands by exocytosis. Endocytosis is basically the opposite process: A small invagination forms in the outer membrane, and the invagination pinches shut, creating a new vesicle that contains extracellular material. En-docytosis is especially common in algae and other microscopic organisms that take in food particles this way, but it is not known how common it is in plants.
Permeability is an important property of membranes. All biological membranes are selectively permeable (also called differentially permeable), meaning that certain substances cross the membrane more easily and rapidly than others (Fig. 5). Because large regions of a membrane are mostly lipid, membranes are more permeable to hydrophobic substances than to anything that carries an electric charge. However, if charged compounds such as inorganic salts, sugars, and amino acids could not enter cells at all, or if they could enter only slowly, cells would starve.
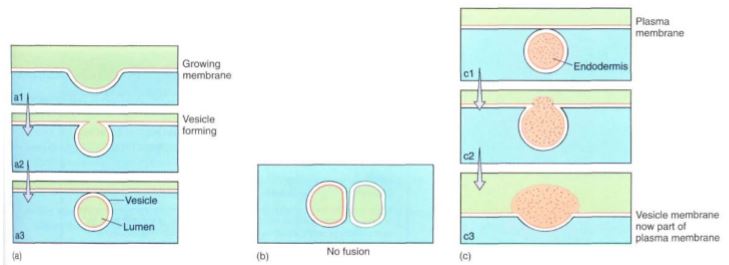
FIGURE 4: Numerous organelles produce small vesicles (a) that move through the cell, either remaining discrete for long periods or fusing with other organelles or vesicles. In order to fuse, two membranes must be quite similar in composition. Here, a single membrane bilayer is shown as a double line, one orange and the other black. (b) Membrane dissimilarity prevents two compartments from fusing; for example, the nucleus does not fuse with mitochondria or chloroplasts. (c) When a vesicle fuses with the membrane at the cell surface, its contents are deposited outside the cell and the vesicle membrane becomes part of the cell surface. The vesicle membrane and the plasma membrane have been drawn with the same colors to indicate their similar composition.
The movement of charged substances is assisted by large intrinsic proteins that span the membrane and act as hydrophilic channels through it; this is facilitated diffusion. Other proteins, called molecular pumps, actually bind to a molecule on one side of the membrane, then by using energy, the protein changes shape and releases the molecule on the other side.
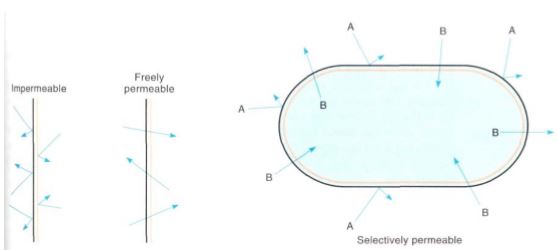
FIGURE 5: An impermeable membrane allows nothing to pass through, whereas a freely permeable membrane stops nothing. A selectively permeable membrane allows certain materials to pass through more readily than others, with the result that chemical concentrations on the two sides differ. This membrane is impermeable to substance A but permeable to B.
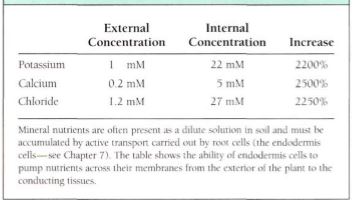
transport, cells accumulate substances until the interior concentration of solute far exceeds the exterior concentration. A pump working in the opposite way can actively transport materials out of the cell (Table1 ). Except for the pumps, the membrane must be impermeable to the molecule; it would do no good to have a molecular pump for a type of molecule that could easily leak back through.
Table 1 :Active Transport in Root of corn
All life depends on the principle of compartmentalization, the formation of many compartments, each specialized for a particular process such as producing a particular substance or using a particular precursor. If all the monomers, polymers, sugars, salts, enzymes, and vitamins of an organism were mixed together, some reactions would occur at random but would not be orderly enough to be considered life. Instead, cells are filled with many compartments-the organelles-each surrounded by its own unique selectively permeable membrane. Because the protein channels and pumps are made under the guidance of the nucleus, the cell can control the numbers and types of channels and pumps it makes and inserts into each type of organelle membrane. Organelles thus have membranes with different permeabilities or pumping capacities, depending on the instructions generated by the nucleus.
A freely permeable membrane, which allows everything to pass through quickly, would be rather useless for a cell, as would an impermeable membrane, one that does not allow anything through at all (Fig. 5; Table 2).
The last important property of membranes is that they are dynamic, constantly changing. Cell membranes are not simply established and left unchanged throughout the life of the cell. Instead, new components are constantly being inserted and old ones removed. If the function of the cell is always the same, the old and new components are similar, but if the cell must change its function, newly inserted components are different from the retracted ones. As the nature of the membrane changes, the nature of the cell changes also.
TABLE 2: Summary of Transmembrane Movement




|
|
|
|
دراسة: حفنة من الجوز يوميا تحميك من سرطان القولون
|
|
|
|
|
|
|
تنشيط أول مفاعل ملح منصهر يستعمل الثوريوم في العالم.. سباق "الأرنب والسلحفاة"
|
|
|
|
|
|
|
الطلبة المشاركون: مسابقة فنِّ الخطابة تمثل فرصة للتنافس الإبداعي وتنمية المهارات
|
|
|
















