آخر المواضيع المضافة

 النبات
الحيوان
الأحياء المجهرية
علم الأمراض
التقانة الإحيائية
التقنية الحيوية المكروبية
التقنية الحياتية النانوية
علم الأجنة
الأحياء الجزيئي
علم وظائف الأعضاء
الغدد
المضادات الحيوية
النبات
الحيوان
الأحياء المجهرية
علم الأمراض
التقانة الإحيائية
التقنية الحيوية المكروبية
التقنية الحياتية النانوية
علم الأجنة
الأحياء الجزيئي
علم وظائف الأعضاء
الغدد
المضادات الحيوية| الارتباط والعبور Likage and crossing over |


|
|
أقرأ أيضاً
التاريخ: 7-11-2019
التاريخ: 2025-03-01
التاريخ: 13-11-2015
التاريخ: 15-11-2015
|
أوضحت نظرية الكروموسومات في الوراثة أن الجينات توجد محمولة على الكروموسومات، أي أن وراثة الجينات لابد وأن تكون عبر انتقال الكروموسومات من جيل إلى جيل.
والمقصود بالارتباط هو أن الجينات الموجودة على نفس الكروموسوم تكون مرتبطة في توارثها بنفس الكروموسوم وبالتالي فهي مرتبطة ببعضها البعض، وفي هذه الحالة فإن هذه الجينات لا تتوزع توزيعاً حراً، لأن التوزيع الحر للجينات ينطبق فقط على الجينات الموجودة على كروموسومات مختلفة. وارتباط الجينات الموجودة على نفس الكروموسوم ببعضها شيء بديهي لأن عدد الجينات بأي كائن حي يفوق كثيراً عدد أزواج الكروموسومات الموجودة به. وعلى سبيل المثال فإن حشرة الدروسوفيلا بها أربعة أزواج فقط من الكروموسومات بينما أوضحت التقديرات التجريبية أن هذه الحشرة تحمل حوالي عشرة الآف جين، ويعني ذلك أن أي كروموسوم لابد وأن يحمل جينات كثيرة جداً. وبما أن الكروموسوم هو وحدة خاصة تنتقل من كل خلية إلى أبنائها، فلابد وأن يصحب معه جميع ما عليه من جينات.
وكما تبين فيما سبق فإن التوزيع الحر للجينات يؤدي في الجيل الثاني إلى ظهور أبناء تشبه أحد الأبوين في صفاتها وتعرف هذه الأبناء بالاتحادات الأبوية (Parental combinations)، وأبناء أخرى لها صفات تظهر مع بعضها باتحادات جديدة (New combinations) تختلف عما كان بالأبوين. والارتباط يعني ميل الاتحادات الأبوية أن تبقى معاً وأن تظهر بنسبة أكبر من النسبة المتوقعة لها على أساس التوزيع الحر للجينات، وبالتالي فإن الاتحادات الجديدة تظهر بنسبة أقل من المتوقع، ويرجع اكتشاف الارتباط إلى كل من "باتسون وبانيت Batson and Punnet" فيما بين عام 1905 وعام 1908 حين لاحظا أن التهجين بين سلالتين من نبات بسلة الزهور "Lathyrus odoratus" يختلفان عن بعضهما في زوجين من الصفات المتفارقة ينتج عنه في الجيل الثاني نسب ظاهرية لا تتفق إطلاقاً مع النسب المتوقعة على أساس التوزيع الحر للجينات. ففي نبات بسلة الزهور يوجد زوجان من الجينات يؤثران على لون الأزهار وشكل حبوب اللقاح وكل زوج من الجينات يوجد بين أليليه علاقة سيادة تام وتنح تام، حيث يسود اللون البنفسجي للأزهار على اللون الأحمر والشكل المستطيل لحبوب اللقاح على الشكل المستدير. أجرى "باتسون وبانيت" تلقيحا بين نبات بنفسجي الأزهار ذي حبوب لقاح طويلة أي يحمل الصفات السائدة النقية (AABB) ونبات أحمر الأزهار مستدير حبوب اللقاح أي متنحي نقي تركيبه الجيني (aabb) وكانت أفراد الجيل الأول كما هو متوقع بنفسجية الأزهار مستطيلة حبوب اللقاح. وطبقا لقانون التوزيع الحر فإن النسب المتوقعة في الجيل الثاني هي 9 : 3 : 3 : 1 بنفسجية الأزهار مستطيلة حبوب اللقاح : حمراء الأزهار مستطيلة حبوب اللقاح : بنفسجية الأزهار مستديرة حبوب اللقاح : حمراء الأزهار مستديرة لحبوب اللقاح على التوالي. ولكن "بتسون وبانيت" حصلا على الأعداد والنسب التالية في الجيل الثاني :

من ذلك يتضح أن تراكيب الصفات الأبوية ظهرت في الجيل الثاني بنسب أكبر من المتوقع حيث ظهرت نباتات بنفسجية مستطيلة حبوب اللقاح ونباتات حمراء الأزهار مستديرة حبوب اللقاح بنسب أعلى من المتوقع لها، بينما ظهرت نباتات ذات اتحادات جديدة أي بنفسجية الأزهار مستديرة حبوب اللقاح وأخرى حمراء الأزهار مستطيلة حبوب اللقاح بنسب أقل من المتوقع لها على أساس التوزيع الحر للجينات.
وعند إجراء التهجين الاختباري لأفراد الجيل الأول وذلك بتلقيحها مع أفراد ذات أزهار حمراء وحبوب لقاح مستديرة نقية حصلا على نفس النتيجة حيث ظهرت نسب للتراكيب الأبوية أكبر كثيراً من النسب المتوقعة بينها كانت نسب الاتحادات الجديدة أقل من المتوقع.
لم يتمكن "باتسون وبانيت" من تقديم تفسير للنسب التي حصلا عليها في تجارب نباتات بسلة الزهور لأنهما لم يفسراها في ضوء الربط بين انتقال جيني لون الأزهار وشكل حبوب اللقاح وبين سلوك الكروموسومات وفسرا نتائجهما هذه إلى وجود تجاذب بين الأليلين السائدين (A,B) وبين الأليلين المتنحيين (a,b) مما أدى إلى بقائهما معاً في جاميتات الجيل الأول وأدى إلى ظهور النسب غير المتوقعة في الجيل الثاني بينما لا تميل الأليلات السائدة والمتنحية إلى الدخول في جاميتة واحدة مما أدى إلى وجود نسب أقل من المتوقعة بالنسبة للاتحادات الجديدة في الجيل الثاني، أطلق على ذلك بالتجاذب و التنافر (Coupling and repulsion )
لم يكن تفسيرهما لهذه الظواهر مرضياً حتى قام العالم "مورجان" وطلبته بتجارب على الدروسوفيلا حصلوا خلاها على نفس النتائج وفسروا نتائجهم إلى ميل الجينات المرتبطة الموجودة على نفس الكروموسوم للبقاء معا في جاميتة واحدة أثناء الانقسام الاختزالي، ويسمى هذا بالارتباط (Linkage). كما بينو أن قوة الارتباط تعتمد على المسافة بين هذه الجينات المرتبطة على الكروموسوم. ولهذا السبب أيضاً فإن نتائج التهجين الاختباري لهذه الجينات المرتبطة تختلف عن النتائج التي يتم الحصول عليها في التهجينات الاختبارية للصفات المندلية نتيجة للتوزيع الحر وهي ما ذكرناه سابقاً بالنسب المعروفة 1:1:1:1 (أي تنحرف هذه النسبة في الجينات المرتبطة).
وفي الدروسوفيلا وعند تلقيح ذباب أصيل للون الجسم الأسود والجناح المختزل مع أخرى برية رمادية وطويلة الجناح فإن جميع أفراد الجيل الأول تكون رمادية اللون طويلة الجناح وعند تلقيح إناث الجيل الأول مع ذكر أسود اللون مختزل الجناح؛ أي تلقيح إناث الجيل الأول مع ذكور ذات صفات متنحية فإن الجيل الأبوي يتألف من فئات أبوية عالية النسبة عما هو متوقع مقارنة بالفئات غير الأبوية الأقل عدداً من المتوقع. ويأتي ذلك بسبب وقوع الأليلين السائدين على كروموسوم بينما الأليلين المتنحيين على الكروموسوم النظير. وبسبب ارتباط الأليلات السائدة مع بعضها والأليلات المتنحية مع بعضها فإن نسب التهجين الاختباري تنحرف عن النسبة المندلية المعروفة، وتقل الفئات غير الأبوية عما هو متوقع. ومن الدراسات العديدة وجد أن هذه النسبة المئوية للاتحادات الجديدة تختلف بين الأزواج المختلفة من الجينات المرتبطة كما وجد أن هذه النسبة دائماً تتراوح بين صفر و 50٪. والشكل الآتي يوضح لنا مثالاً للاتحادات الأبوية والاتحادات الجديدة التي ظهرت في أحد تجارب "مورجان" وأنها تنحرف عن النسب المتوقعة.

شكل (1) احد تجارب العالم مورجان حيث يبين النسل الناتج من التهجين الاختباري لاحد افراد الجيل الأول والذي يحمل الصفات السائدة الهجينة بالنسبة للون الجسم الرمادي والاجنحة الطبيعية مع فرد يحمل الصفات المتنحية ( اسود مختزل الجناح )
ونتيجة لذلك وبعد دراسة حالات متعددة في الدروسوفيلا، أدرك «مورجان» أن التجاذب والتنافر هما حالتان لظاهرة واحدة وهي ما أطلق عليها اسم الارتباط (Linkage) ينتج عنها أن الاتحادات الأبوية دائماً تظهر بنسب أعلى من المتوقع، نتيجة لميل الجينات المرتبطة - أي الموجودة على كروموسوم واحد - إلى الانتقال كتلة واحدة، أي تكون مرتبطة ببعضها. كما فسر ظهور الاتحادات الجديدة بين الجينات المرتبطة إلى ظاهرة أخرى أطلق عليها اسم العبور (Crossing over)، وتتلخص في أنه يحدث تبادل مادي بين فردي كل زوج من الكروموسومات النظيرة أثناء الانقسام الاختزالي وتكوين الجاميتات. وقد كانت جميع البيانات الوراثية والمشاهدات السيتولوجية التي حصل عليها "مورجان" و مساعدوه من دراساتهم على الدروسو فيلا أكبر دعامة لهذا التفسير.
ويمكن تفسير ظاهرة العبور Crossing over" من تتبع سلوك الكروموسومات أثناء الانقسام الميوزي حيث يلاحظ حدوث ظاهرة تفسر انفصال الجينات المرتبطة. فأثناء الطور الضام نجد أن كل كروموسوم ينشق طولياً إلى كروماتيدتين ويتلاصق كل كروموسومين متماثلين لتكوين الوحدة الكروموسومية الثنائية (Bivalent) المكونة من أربع كروماتيدات. وفي نفس الوقت تحدث عملية تبادل بين كروماتيدتين غير شقيقتين من الكروموسومين المتماثلين عند نقطة أو أكثر فتتكون بذلك الكيازمات (Chiasmata) التي تشاهد بوضوح عند الطور التشتتي وبذلك يحدث انتقال جينات من كروموسوم إلى مثيله.
ونتيجة لهذه العملية، أي بعد عملية العبور نجد أنه بجانب التراكيب الأبوية يوجد تراكيب جديدة. وقد اعتبر مورجان أن هذا التعليل هو التفسير الصحيح لعملية انتقال الجينات من كروموسوم لآخر وذلك يفسر ظهور أفراد ذات صفات غير أبوية أي اتحادات جديدة في النسل الناتج من التلقيح الاختباري. ويتوقف معدل العبور أو نسبة العبور وقوة الارتباط على المسافة بين الجينات، فإذا كانت هذه المسافة بين الجينين كبيرة كان احتمال حدوث عبور بينهما كبير، وإذا تقارب الجينان كان الارتباط بينهما قوياً، فكلما زادت المسافة بين موقعي الجينين كلما زاد احتمال تكوين الكيازمات وبالتالي زادت نسبة الحالات التي ينتقل فيها الجين من كروموسوم لآخر أي العبور. ويمكن تحديد نسبة العبور أو معدل الارتباطات الجديدة بين الجينات بعدد الافراد التي تمثل الاتحادات الجديدة منسوباً الى العدد الكلي للأفراد أي :
نسبة العبور = عدد الافراد التي بها اتحادات جديدة / المجموع الكلي للافراد × 100
ويطلق على الافراد ذات الاتجاهات الجديدة فئات عبورية بينما يطلق على الاتحادات الابوية فئات غير عبورية اوارتباطية.
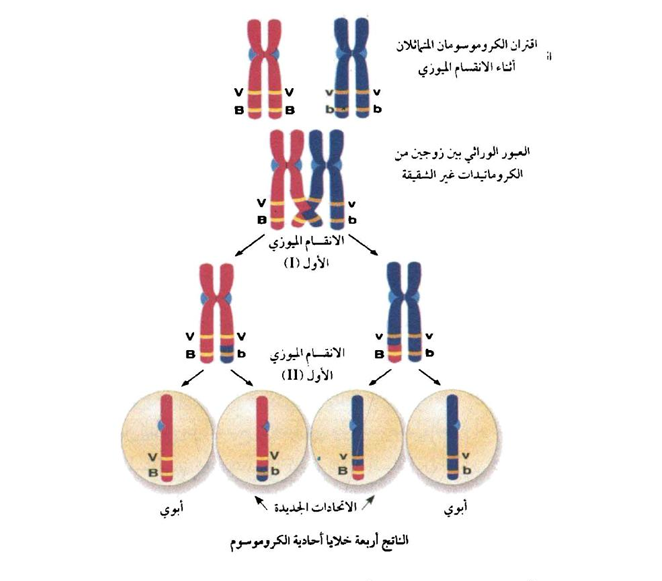
شكل (2) يوضح كيفية حدوث العبور اثناء الانقسام الاختزالي وكيفية تكوين الاتحادات الجديدة

شكل (3) يبين الكيازمات المتكونة اثناء عملية العبور الوراثي



|
|
|
|
دراسة تكشف "مفاجأة" غير سارة تتعلق ببدائل السكر
|
|
|
|
|
|
|
أدوات لا تتركها أبدًا في سيارتك خلال الصيف!
|
|
|
|
|
|
|
العتبة العباسية المقدسة تؤكد الحاجة لفنّ الخطابة في مواجهة تأثيرات الخطابات الإعلامية المعاصرة
|
|
|