
آخر المواضيع المضافة

 النبات
الحيوان
الأحياء المجهرية
علم الأمراض
التقانة الإحيائية
التقنية الحيوية المكروبية
التقنية الحياتية النانوية
علم الأجنة
الأحياء الجزيئي
علم وظائف الأعضاء
الغدد
المضادات الحيوية
النبات
الحيوان
الأحياء المجهرية
علم الأمراض
التقانة الإحيائية
التقنية الحيوية المكروبية
التقنية الحياتية النانوية
علم الأجنة
الأحياء الجزيئي
علم وظائف الأعضاء
الغدد
المضادات الحيوية| Depurination |


|
|
Read More
Date: 5-3-2021
Date: 3-1-2016
Date: 10-3-2021
|
Depurination
An abasic site one lacking a nucleotide base, is created at a site in DNA when the glycosidic bond connecting a purine base to the deoxyribose sugar is cleaved, leaving the phosphodiester backbone of the DNA intact (Fig. 1). Depurination occurs at an estimated rate of 104 depurination per mammalian cell per day (1), often through spontaneous hydrolysis. Other processes in the cell that lead to depurination are enzymatic removal of altered bases by specific glycosylases or through chemical modifications of purine bases (primarily alkylation at N7) that labilize the glycosidic bond. There is good evidence that depurination plays a major role in spontaneous mutagenic events (2). Although apurinic sites inhibit chain elongation by DNA polymerases, bypass must occur because there is enhanced incorporation of nucleotides that are noncomplementary to the template bases. The rate of misincorporation varies with different polynucleotide complexes, but the frequency of misincorporation is directly proportional to the extent of depurination (2).
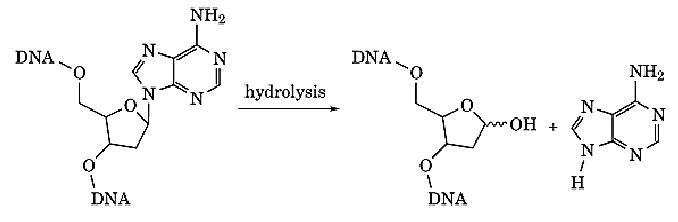
Figure 1. Formation of an abasic site in DNA depurination.
Weymouth and Loeb (3) and Kunkel et al. (4) studied the fidelity of DNA polymerases in copying natural DNA templates and the mutagenic implications of the misincorporation events. They measured revertants generated by misincorporation in am3 bacteriophage fX174, a mutant containing a single base-pair substitution mutation. These studies confirmed that the copying of depurinated am3 fX174 DNA templates by purified DNA polymerases is mutagenic, and that the frequency of revertants correlates positively with the ability of different polymerases to copy past apurinic sites. Eukaryotic DNA polymerases are error-prone and read past apurinic sites at a high frequency.
Although direct mutagenicity is readily envisaged following error-prone read through of an apurinic site, it also appears that many mutations resulting from depurination in prokaryotes are SOS-response-dependent (5). Miller and Low (6) examined more than 600 independent spontaneous mutations in the lacI gene of SOS-induced E. coli. Most were specifically G.C → T.A, and to a lesser extent A.T → T.A, transversions and occurred primarily at certain sites. These transversion mutations are thought to have arisen from the insertion of dAMP opposite purines or apurinic sites, possibly through the SOS-induction process altering the specificity of the DNA replication complex to favor misincorporation of dAMP opposite purines. Miller and Low suggested that many spontaneous mutations may result from replication past cryptic apurinic sites. There are also several lines of evidence that apurinic sites are intermediates in mutagenesis by chemicals that form bulky DNA adducts (2).
References
1. T. Lindahl and B. Nyberg (1972) Biochemistry 11, 3610–3618.
2. L. A. Loeb (1985) Cell 40, 483–484.
3. L. A. Weymouth and L. A. Loeb (1978) Proc. Natl. Acad. Sci. USA 75, 1924–1928.
4. T. A. Kunkel, R. M. Schaaper, and L. A. Loeb (1983) Biochemistry 22, 2378–2384.
5. T. A. Kunkel (1984) Proc. Natl. Acad. Sci. USA 81, 1494–1498.
6. J. H. Miller and K. B. Low (1984) Cell 37, 675–682.



|
|
|
|
دخلت غرفة فنسيت ماذا تريد من داخلها.. خبير يفسر الحالة
|
|
|
|
|
|
|
ثورة طبية.. ابتكار أصغر جهاز لتنظيم ضربات القلب في العالم
|
|
|
|
|
|
|
العتبة العباسية المقدسة تستعد لإطلاق الحفل المركزي لتخرج طلبة الجامعات العراقية
|
|
|
















