
آخر المواضيع المضافة

 النبات
الحيوان
الأحياء المجهرية
علم الأمراض
التقانة الإحيائية
التقنية الحيوية المكروبية
التقنية الحياتية النانوية
علم الأجنة
الأحياء الجزيئي
علم وظائف الأعضاء
الغدد
المضادات الحيوية
النبات
الحيوان
الأحياء المجهرية
علم الأمراض
التقانة الإحيائية
التقنية الحيوية المكروبية
التقنية الحياتية النانوية
علم الأجنة
الأحياء الجزيئي
علم وظائف الأعضاء
الغدد
المضادات الحيوية| Chloramphenicol |


|
|
Read More
Date: 1-11-2020
Date: 28-4-2021
Date: 22-12-2015
|
Chloramphenicol
Chloramphenicol is a bacteriostatic agent that inhibits the growth of many species of Gram-positive and Gram-negative bacteria; it was the first broad-spectrum antibiotic to be used clinically. Originally obtained from cultures of the soil bacterium Streptomyces venezuelae, chloramphenicol inhibits bacterial protein biosynthesis by interfering with the intrinsic catalytic activity of the peptidyl-transferase of the ribosome during the elongation phase of translation.
Eukaryotic cells are generally not affected by chloramphenicol. Nevertheless, the clinical application of this drug for the treatment of systemic infections has been severely curtailed, primarily because of the haematotoxicity associated with its use. Chloramphenicol continues to be used topically, particularly in the treatment of eye infections. The emergence of chloramphenicol-resistant bacteria, apparently in response to the selective pressure exerted by the drug, has also restricted the use of chloramphenicol in the treatment of bacterial infections.
Chloramphenicol resistance is frequently encountered in many genera of bacteria. Several mechanisms of resistance have been described: (i) modification of the target, the bacterial ribosome; (ii) alteration in the permeability of the bacterial cell, leading to low intracellular antibiotic concentrations; and (iii) enzymatic modification of the antibiotic. Resistance most commonly occurs as the result of inactivation of chloramphenicol by chloramphenicol acetyltransferase (CAT). This enzyme is of value to molecular biologists, because it can be assayed with specificity and sensitivity.
Chloramphenicol is used today in molecular biology primarily as a component of selective media for genetic studies, to distinguish chloramphenicol-resistant from chloramphenicol-susceptible bacteria. Chloramphenicol is also an essential component of CAT assays, which are used to study gene regulation, generally in eukaryotic cells. Finally, chloramphenicol provides a tool with which to investigate the peptidyl-transferase center of the bacterial ribosome.
1. Chemistry
Chloramphenicol [D(–)-threo-2-dichloroacetamido-1-p-nitrophenyl-1,3-propanediol; Fig. 1] has two asymmetric carbon atoms. Of the four stereoisomers, only one, the D-threo isomer, has antibacterial activity. This compound can be produced on an industrial scale by chemical synthesis or by fermentation. Chloramphenicol is poorly soluble in water, but dissolves readily in organic solvents (eg, methanol).
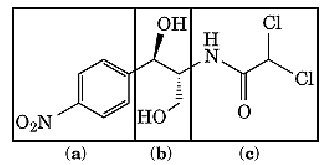
Figure 1. Chemical structure of chloramphenicol [D(–)-threo-2-dichloroacetamido-1-p-nitrophenyl-1,3-propanediol].
Derivatives having a wide range of para substituents are active; therefore, this part of the molecule is not involved in specific drug–target interactions. The propanediol moiety, the site of the D-threo configuration, is essential for activity. Acetylation of one or both hydroxyl groups inactivates the drug. Removal of the dichloroacetamido side chain virtually eliminates biological activity, although substitution by a number of groups does not render the drug inactive. Some substituents reduce activity against intact bacteria, but enhance the inhibition of cell-free protein synthesis (1). Apparently, these derivatives are not taken up by the bacterial cell as efficiently as the parent compound.
2. Antibacterial Spectrum
Chloramphenicol is a broad-spectrum antibiotic, active against archaebacteria and both gram-positive and gram-negative bacteria. Organisms that fall within its spectrum include Bacillus anthracis, Brucella abortus, Corynebacterium diphtheriae, Escherichia coli, Hemophilus influenzae, Klebsiella pneumoniae, Neisseria meningitidis, Salmonella spp., Serratia marcescens, Staphylococcus aureus, Streptococcus spp., and so on (2).
Chloramphenicol inhibits translation in bacterial cells, intact chloroplasts, and intact mitochondria. Some of the toxic side effects associated with chloramphenicol may arise from the effect of the drug on mitochondrial metabolism. Translation by mammalian cells, yeast, and protozoa is not inhibited by chloramphenicol.
3. Mode of Action
Chloramphenicol inhibits bacterial peptidyl-transferase, the enzymatic reaction by which one more amino acid is added to the growing polypeptide chain. During the elongation phase of protein biosynthesis in bacteria, chloramphenicol “freezes” polysomes on the messenger RNA, thereby fixing the peptidyl-tRNA to the ribosomes (3). Chloramphenicol competes for binding to the 50S subunit of the bacterial ribosome with molecules that specifically bind to the A site of the peptidyl-transferase center (aminoacyl-tRNA and its analogues, eg, CACCA-Leu) and is, therefore, thought to bind next to, or within, the A site of the peptidyl-transferase center (4). In contrast, no competition is observed with substrates that bind at the P site (peptidyl-tRNA analogues, eg, CACCA-LeuAc or AcPhe-tRNA). Furthermore, neither the dinucleotide ApC nor amino acids block the binding of chloramphenicol, whereas both puromycin and the dinucleotide CpA do (5). Thus, the drug presumably binds in the same region of the A site as the universally conserved CCA part of aminoacyl-tRNA.
The peptidyl-transferase assay in which peptide bonds are formed between a peptidyl-tRNA and puromycin on the 50S subunit (the “puromycin reaction”) has been used to unravel the mechanism of action of chloramphenicol (6) (Fig. 2). Chloramphenicol blocks this reaction, provided that the donor peptidyl-tRNA, bound at the P site, is short (eg, AcPhe-tRNA). In this case, chloramphenicol appears to disturb the correct positioning of the nascent peptide during its synthesis, thereby causing the premature release of the peptidyl-tRNA from the ribosome. Chloramphenicol does not, however, affect the puromycin reaction when a larger peptidyl-tRNA—for example, Ac(Phe)2-tRNA—is used as donor (7). It is likely, therefore, that chloramphenicol interferes with the binding of the terminal CCA of the aminoacyl-tRNA at the A site and also interferes with the entry of the nascent polypeptide into the “tunnel” that normally guides it away from the peptidyl-transferase center.
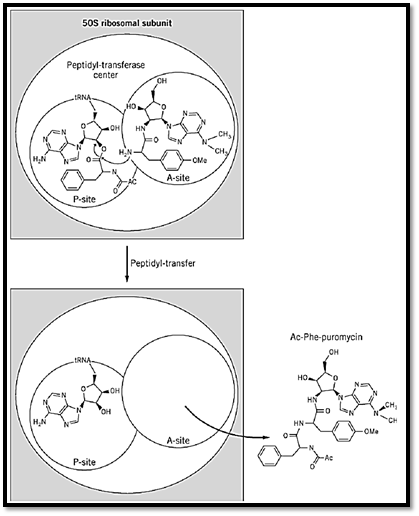
Figure 2. The puromycin reaction. A peptidyl-tRNA analogue [eg, Ac[14C]Phe-tRNA) bound to the ribosomal P site of 50S ribosomal subunit reacts with puromycin (an aminoacyl-tRNA analogue) bound to the A site of the peptidyl-transferase center. The resulting peptidyl-puromycin (eg, Ac[14C]Phe-puromycin) is falling of the 50S subunit and can be separated from the substrate (Ac[14C]Phe-tRNA) by extraction.
Chloramphenicol inhibits protein synthesis in bacterial extracts with varying potencies depending on the template employed. In particular, synthesis promoted by poly(U) is markedly more resistant to chloramphenicol than that promoted by poly(C) or poly (A) (8). Synthesis occurring with a natural mRNA (eg, MS2 phage RNA) is usually very sensitive to chloramphenicol. The precise reason for the template-dependence is not known. Chloramphenicol may inhibit poly(U)-dependent poly(Phe) synthesis less than it does the synthesis of other polypeptides because the structures of D-threo chloramphenicol and L-phenylalanine are similar (Fig. 3). Competition between these molecules during poly(Phe) synthesis may involve steric hindrance between their phenyl groups and, consequently, chloramphenicol may be chased from the ribosome. Nevertheless, this does not explain why the same template dependence is also observed with several other peptidyl-transferase inhibitors that differ chemically from chloramphenicol and, in particular, that do not have a phenyl group (e.g, the group B streptogramins and the macrolides). Further experiments are needed to elucidate this phenomenon in greater detail.
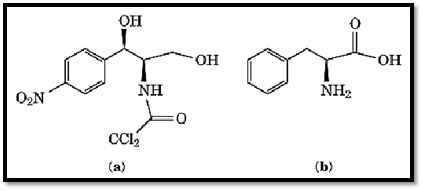
Figure 3. Structural similarity of D(–)-threo chloramphenicol and L-phenylalanine. Competition between these molecules during poly(Phe) synthesis may involve steric hindrance between their phenyl groups.
4. Ribosomal Binding Site
Chloramphenicol has a single high-affinity binding site located on the 50S ribosomal subunit (Kd = 2 × 10–6 M), as well as one or more low-affinity sites, at least one of which is located on the 30S subunit (Kd = 2 × 10–4 M) (9). A number of 50S proteins were labeled in affinity-labeling experiments, including L1, L2, L11, L16, L19, and L27 (10). Most of these proteins are located in or next to the peptidyl-transferase center of the ribosome, and several can be affinity-labeled with other antibiotics that interact with this center (erythromycin, puromycin, lincosamides, streptogramins).
That the peptidyl-transferase domain is the primary binding site of chloramphenicol was confirmed by RNA footprinting experiments, in which bases of 23S rRNA were compared in antibiotic-free or chloramphenicol-bound ribosomes for their reactivity to chemical probes. Chloramphenicol protected bases A2059, A2062, A2451, and G2505 and enhanced the reactivity of A2058 (11). Protection of A2070 and U2506 has also been observed (12). All these nucleotides are located in the central loop of domain V of 23S rRNA (Fig. 4). Additionally, several mutations of RNA that confer resistance to chloramphenicol have been identified in the same domain (Fig. 4; see references in Ref. 12).
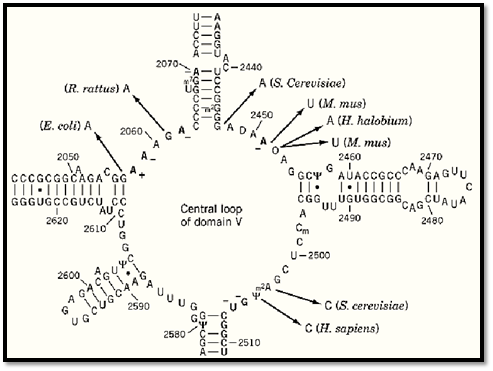
Figure 4. Interaction of chloramphenicol with the central loop of domain V of 23S rRNA. The secondary structure of the central loop of domain V in E. coli 23S rRNA is derived from phylogenetic sequence comparisons. Differences in the accessibility of bases to chemical reagents in the presence and absence of chloramphenicol (footprints) are shown in red (–, protection of base; +, enhanced activity). Mutations leading to chloramphenicol resistance are indicated by arrows, and the corresponding organisms are indicated in brackets. All of the eukaryotic mutations were characterized for mitochondrial rRNA. (Figure derived from Refs. 11 and 12.)
5. Cold-Shock Induction
Shifting growing E. coli from 37°C to 20°C inhibits cellular growth and shuts down protein biosynthesis. To compensate for the stress produced by the temperature shift, the levels of (p)ppGpp drop and the synthesis of a subset of proteins is induced; this induction is known as cold-shock. Certain inhibitors of protein synthesis, including chloramphenicol, specifically induce the synthesis of the cold-shock proteins in the absence of a temperature shift (13). Ribosomal inhibitors have, in fact, been classified according to their ability to elicit a cold-shock- or a heat-shock-like response. Inhibitors in whose presence the A site of the peptidyl-transferase center is occupied, either by the antibiotic or by an aminoacyl-tRNA (chloramphenicol, erythromycin, fusidic acid, spiramycin, tetracycline), produce a cold-shock-like response. In contrast, inhibitors in whose presence the A site of the peptidyl-transferase remains empty (kanamycin, streptomycin) produce a heat-shock-like response.
6. Chloramphenicol Resistance
6.1. Modification of the Bacterial Ribosome
Several mutations in 23S rRNA confer resistance to chloramphenicol. The residues at which these mutations occur (residues G2057, G2061, G2447, A2451, C2452, A2503, U2504; E. coli numbering) are located in the peptidyl-transferase loop of 23S rRNA (Fig. 4), which is in accordance with their proposed binding site.
6.2. Alteration in Permeability to Chloramphenicol
Protein synthesis in S. venezuelae cells that have begun to produce chloramphenicol becomes insensitive to the antibiotic. There is no modification of the ribosome in these cells; instead, the antibiotic is excluded from the cytoplasm during biosynthesis. The amino acid sequence deduced from the gene conferring resistance (cml) is markedly similar to those of chloramphenicol-resistance genes from Streptomyces lividans, Rhodococcus fascias, and other bacteria. These polypeptides contain 12 hydrophobic regions characteristic of membrane-associated transport proteins and are related to a known family of efflux proteins (14).
6.3. Enzymatic Modification of Chloramphenicol
The most common mechanism of chloramphenicol resistance is inactivation by the enzyme chloramphenicol 3′-O-acetyltransferase (CAT), encoded by the cat gene. The biological and immunological properties of three types of CAT (I, II, III) have been described. The structure of CAT III has been determined by X-ray crystallography, as well as its tertiary complex with chloramphenicol and CoA (15). The enzyme is a trimer; its active sites lie at each of the interfaces between adjacent subunits. CAT catalyzes the transfer of an acetyl moiety from acetyl-CoA to the 3′-hydroxyl of chloramphenicol. The initial product of the reaction, 3-acetyl chloramphenicol, can undergo nonenzymatic intramolecular rearrangement to 1-acetyl chloramphenicol. The latter compound is a substrate for a second round of enzymatic acetylation at the 3-hydroxyl, which yields 1,3-diacetyl chloramphenicol as the final product (Fig. 5). Neither the monoacetylated nor the diacetylated derivative binds to the bacterial ribosome; both, therefore, are devoid of antibiotic activity.
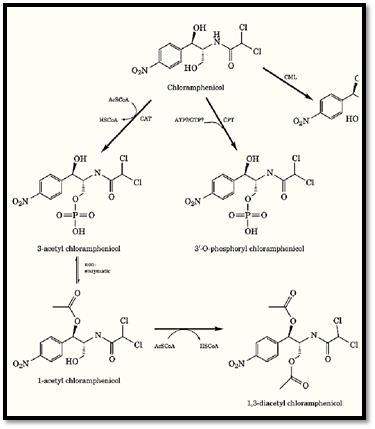
Figure 5. Enzymatic inactivation of chloramphenicol. CAT, chloramphenicol 3′-O-acetyltransferase; CPT, chloramphen CML, chloramphenicol hydrolase.
Recently, a second chloramphenicol 3′-O-modifying gene was discovered (16). This enzyme, chloramphenicol 3′-O-phosphotransferase (CPT) phosphorylates chloramphenicol, yielding 3′-O-phosphochloramphenicol, which binds only very weakly to ribosomes and thus does not inhibit translation.
Chloramphenicol is also inactivated by chloramphenicol hydrolase, which hydrolyzes the amide bond (17) (Fig. 5). This enzyme, encoded by the cml gene, was first isolated from the mycelium of a chloramphenicol-producing strain of Streptomyces. A similar enzyme has been obtained from several other bacteria, including E. coli.
6.4. Induction of Chloramphenicol Resistance by Translational Attenuation Chloramphenicol resistance by enzymatic modification in both Gram-positive and Gram-negative bacteria can be either constitutive or inducible. What is unusual about inducible chloramphenicol resistance is that it occurs by translational attenuation, rather than by transcriptional attenuation. The ribosome-binding site for the resistance determinant is sequestered in a secondary “hairpin” structure situated within the encoding mRNA; a short, translated open reading frame, termed the leader, lies upstream of this structure. In the absence of the antibiotic, the secondary structure is maintained, the ribosome does not translate the mRNA that constitutes the hairpin, and the resistance gene is not translated. In the presence of the antibiotic, the ribosome stalls at the leader sequence and the secondary structure relaxes, allowing the cat (or cml) determinant to be translated (18) (Fig. 6).
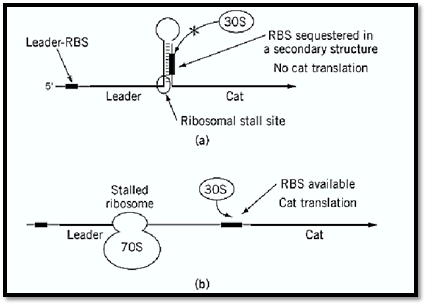
Figure 6. Chloramphenicol resistance by translational attenuation. RBS, ribosomal binding site; 30S, small ribosomal subunit; 70S, entire bacterial ribosome.
7. The Use of CAT in Molecular Biology
CAT is a gene product that can be assayed with specificity and sensitivity, and consequently it is of considerable value to molecular biologists. When the cat gene is fused to heterologous regulatory signals, gene expression can be determined by measuring the CAT activity. Eukaryotes lack endogenous CAT activity, and the amount of CAT activity correlates well with the amount of mRNA transcribed, so the CAT assay has proved to be a powerful tool for investigating and quantifying gene expression in eukaryotic cells. Conventional CAT assays involve the incubation of cell extracts with radioactively labeled chloramphenicol, chromatographic separation of the reaction products, and autoradiography. The recent use of fluorescent, rather than radioactive, substrates has dramatically speeded up and simplified the CAT assay (19).
References
1. O. Pongs (1979) In Antibiotics Vol. 1 (F. H. Hahn ed.), Springer-Verlag, Heidelberg, Germany, pp. 26–56.
2. A. A. Yunis (1988) Annu. Rev. Pharmacol. Toxicol. 28, 83–100.
3. E. Cundliffe and K. McQuillen (1967) J. Mol. Biol. 30, 137–143.
4. D. Nierhaus and K. H. Nierhaus (1973) Proc. Natl. Acad. Sci. USA 70, 2224–2228.
5. M. L. Celma, R. E. Monro, and D. Vazquez (1971) FEBS Lett. 13, 247–251.
6.R. R. Traut and R. E. Monro (1964) J. Mol. Biol. 10, 63–72.
7. H.-J. Rheinberger and K. H. Nierhaus (1990) Eur. J. Biochem. 193, 643–650.
8. Z. Kucan and F. Lipmann (1964) J. Biol. Chem. 239, 516–520.
9. J. L. Lessard and S. Pestka (1972) J. Biol. Chem. 247, 6909.
10. B. S. Cooperman, C. J. Weitzmann, and C. L. Fernandez (1990) In The Ribosome: Structure,
Function, & Evolution (W. E. Hill et al., eds.), American Society for Microbiology, Washington, D.C., pp. 491–501.
11. D. Moazed and H. F. Noller (1987) Biochimie 69, 879–884.
12. C. Rodriguez et al. (1995) J. Mol. Biol. 247, 224–235.
13. P. A. Lagosky and F. N. Chang (1981) J. Biol. Chem. 256, 11651–11656.
14. H. Nikaido and M. H. Sailer Jr. (1992) Science 258, 936–942.
15. A. G. W. Leslie (1990) J. Mol. Biol. 213, 167.
16. R. H. Mosher et al. (1995) J. Biol. Chem. 270, 27000–27006.
17. V. S. Malik and L. C. Viking (1971) Can. J. Microbiol. 17, 1287–1290.
18. P. S. Lovett (1996) Gene 179, 157–162.
19. C. K. Lefevre et al (1995) Biotechniques 19, 488–493.



|
|
|
|
تفوقت في الاختبار على الجميع.. فاكهة "خارقة" في عالم التغذية
|
|
|
|
|
|
|
أمين عام أوبك: النفط الخام والغاز الطبيعي "هبة من الله"
|
|
|
|
|
|
|
المجمع العلمي ينظّم ندوة حوارية حول مفهوم العولمة الرقمية في بابل
|
|
|
















