
آخر المواضيع المضافة

 النبات
الحيوان
الأحياء المجهرية
علم الأمراض
التقانة الإحيائية
التقنية الحيوية المكروبية
التقنية الحياتية النانوية
علم الأجنة
الأحياء الجزيئي
علم وظائف الأعضاء
الغدد
المضادات الحيوية
النبات
الحيوان
الأحياء المجهرية
علم الأمراض
التقانة الإحيائية
التقنية الحيوية المكروبية
التقنية الحياتية النانوية
علم الأجنة
الأحياء الجزيئي
علم وظائف الأعضاء
الغدد
المضادات الحيوية| Acentric Fragment |


|
|
Read More
Date: 25-4-2016
Date: 7-3-2021
Date: 3-5-2021
|
Acentric Fragment
Fragments of chromosomes that lack centromeres are described as acentric fragments (Fig. 1). They are formed as a result of chromosomal damage. Lacking a centromere, there is no means of attachment to the mitotic spindle, which leads to a failure to segregate replicated chromatids in the acentric fragments to the daughter cells during cell division. Therefore, the acentric fragments are normally lost from most cells with progressive cell cycles. The breakage of the chromosome that separates the acentric fragment from the remainder of the chromosome containing the centromere can be a result of either inherent chromosomal fragility, radiation damage, or defective DNA repair mechanisms.
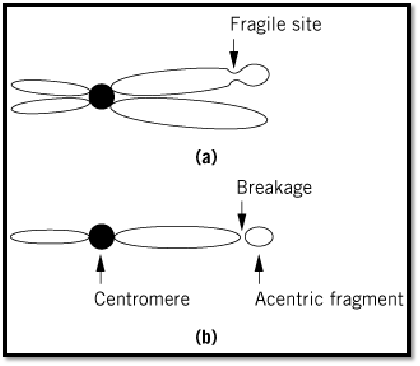
Figure 1. (a) A mitotic chromosome is shown with a fragile site, indicated. (b) Mitosis can create forces that break the chromosome at the fragile site, creating a residual chromosome that contains the centromere and an acentric fragment as indicated.
Chromosome fragile sites lead to acentric fragments in humans. The majority of recurrent cancer chromosome breakpoints are found at fragile sites (1). These breakpoints may promote oncogenic rearrangements and translocations. Chromosomal fragile sites can be induced by growing cells under abnormal conditions, by the addition of drugs or chemicals, and by infection with viruses. Fragility seems to be a consequence of incomplete compaction of the chromatin. A more open and accessible chromatin structure might explain the tendency of these sites to break, to recombine with other chromosomal regions, and to be sites of viral integration (2). A fragile site associated with a form of inherited mental retardation known as fragile X mental retardation has been characterized in some detail (5). A gene called FMR-1 (fragile X mental retardation gene 1) is present in the fragile site. This gene contains within it a trinucleotide repeat of CGG. A normal individual has between 6 and 50 of these trinucleotide repeats. Individuals with fragile X syndrome contain expansions of these repeat sequences such that they have several hundred CGG triplets. This expansion leads to chromosome fragility, to inactivation of FMR-1 gene expression, and to methylation of regulatory DNA within the gene. The expansion of the CGG trinucleotide sequences is associated with the assembly of an aberrant chromatin structure (3).
Defective chromosomal repair mechanisms lead to a much higher number of chromosomal breaks and translocations than found in normal cells. This high degree of chromosomal damage can eventually lead to malignancy. For example, in Bloom's syndrome there is a very high rate of sister chromatid exchange, and in Fanconi's anemia and ataxia telangiectasia there is a high incidence of chromosomal breakage and rejoining, leading to multicentric chromosomes and acentric fragments. Individuals with these syndromes are especially sensitive to environmental insults, such as chemical mutagens and ionizing radiation, which further increase chromosomal damage (4). Detection of acentric fragments is diagnostic of serious abnormalities in chromosomal metabolism.
References
1. J.J. Yunis and A.L. Soreng, Science 226, 1199–1204 (1984).
2. A.D. Bailey, Z. Li, T. Pavelitz, and A.M. Weiner, Mol. Cell. Biol. 15, 6246–6255 (1995).
3. R.S. Hansen et al., Cell 73, 1403–1409 (1993).
4. C.J. Vessey, C.J. Norbury, I.D. Hickson, Prog Nucleic Acid Res Mol Biol 63, 189–221 (1999).
5. P. Jin and S.T. Warren, Hum. Mol. Genet. 9, 901–908 (2000).



|
|
|
|
علامات بسيطة في جسدك قد تنذر بمرض "قاتل"
|
|
|
|
|
|
|
أول صور ثلاثية الأبعاد للغدة الزعترية البشرية
|
|
|
|
|
|
|
مكتبة أمّ البنين النسويّة تصدر العدد 212 من مجلّة رياض الزهراء (عليها السلام)
|
|
|
















