تنتمي الحشرات إلى تحت شعبة الحشرات (Sub-Phylum : Hexapoda (Insecta والتي تضم 29 رتبة Orders، إلا أن ثمانية رتب منها فقط تحوي ناقلات لفايروسات النبات وجميع هذه الرتب الثمانية تضم حشرات نباتية التغذية، (الجدول 1) وتعد رتبة متساوية الأجنحة Homoptera هي الرتبة الحشرية الأهم في نقل الفايروسات لأنها تضم أهم الناقلات الحشرية تليها رتبة غمدية الأجنحة Coleoptera ثم رتبة هدبية الأجنحة (الثربس) Thysanoptera، كما سجلت أنواع قليلة ناقلة تعود لرتب جلدية الأجنحة Dermaptera ومستقيمة الأجنحة Orthoptera وحرشفية الأجنحة Lepidoptera وثنائية الأجنحة Diptera ولكنها قليلة الأهمية في نقل الفايروسات وأن العلاقة بينهما هي علاقة ميكانيكية بسيطة غالبا، (الشكلين 1 و 2). وتعد الحشرات ذات أجزاء الفم الثاقبة الماصة Piercing-sucking mouthparts أهم الحشرات الناقلة لفايروسات النبات والمسؤولة عن نقل ونشر العديد من أنواع الفايروسات الخطرة المهمة اقتصاديا وأن العلاقة بينهما هي من أعقد العلاقات الحيوية المتخصصة بين الطفيليات وناقلاتها حيث وصل الأمر إلى تضاعف بعض الأنواع الفايروسية في ناقلاتها الحشرية مما يعني أن هذه الفايروسات هي مزدوجة التطفل على النبات والحشرة معا، وتتأثر العلاقة المعقدة بين الحشرات والفايروسات بالتداخل بين الفايروس والنبات العائل والحشرة الناقلة والظروف البيئية.
الجدول (1): المجاميع التصنيفية لناقلات فايروسات النبات
تقسم الفايروسات النباتية عموما إلى مجموعتين أساسيتين حسب علاقتها مع ناقلاتها الحشرية وهما (1) الفايروسات المحمولة داخليا في جسم الحشرة Internally-borne Viruses وتشمل ثلاثة مجاميع من الفايروسات هي الفايروسات شبه الباقية Semi-persistent viruses والفايروسات الدوارة Circulative viruse والفايروسات المتكاثرة Propagative Viruses (2) الفايروسات المحمولة خارجيا على جسم الحشرة Externally-borne Viruses والتي تحمل حصرا على الجزء الأمامي من أجزاء الفم (الرمح) وتشمل "الفايروسات غير الباقية" Non-persistent Viruses والتي يطلق عليها ايضا الفايروسات المحمولة بالرمح .Stylet-borne Viruses
يطلق على الفايروسات شبه الباقية والفايروسات غير الباقية مصطلح "الفايروسات غير الدوارة" Non-Circulative Viruses لأنها لا تدخل إلى دم الحشرة وتدور فيه. هناك ظاهرة مثيرة الاهتمام في علاقة الفايروسات بناقلاتها عموما وبضمنها الحشرات وهي أن الفايروسات المنقولة بمجموعة معينة من الناقلات لا تنقل بمجموعة أخرى ويحصل هذا التخصص في النقل على مستوى مجموعة الناقل Vector type أي حشرات أو عناكب أو نيماتودا الخ، او يحصل على مستوى عائلة او على مستوى الجنس او النوع واحيانا على مستوى النمط الحيوي للناقل Biotype.

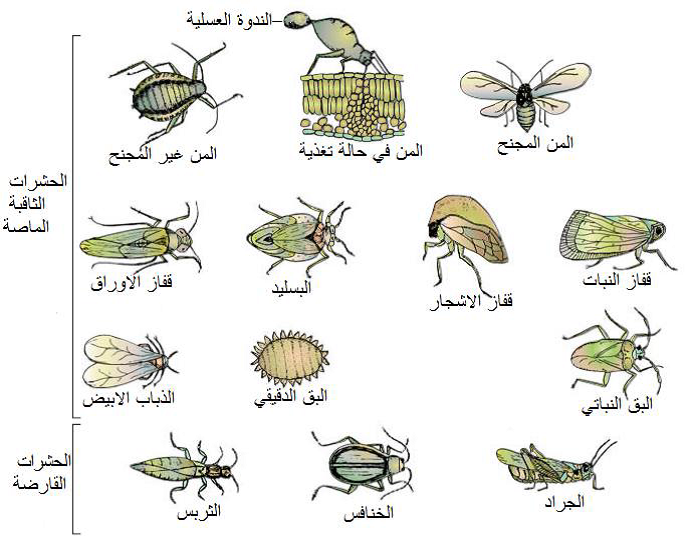
الشكل (1): بعض انواع الحشرات الناقلة للفايروسات.
الشكل مقتبس من Carter و Saunders (2007).
الشكل (2): رسوم توضيحية لأنواع الحشرات الثاقبة الماصة والقارضة الناقلة لفايروسات النبات.
المصدر: Agrios 2005))
(1) النقل بحشرات المن
حشرات المن Aphids هي أهم ناقلات فايروسات النبات إذ بلغ عدد أنواع المن المسجلة منها ما يقرب من 3740 نوعا وهي تنتمي تصنيفيا إلى فوق العائلة Super family: Aphidoidea تضم ثلاث عوائل وتعد العائلة Aphidiidae هي أهم العوائل التي تضم أغلب أنواع المن الناقلة للفايروسات أما العائلة Phylloxeridae فتضم 60 نوعا من المن ولكنها قليلة الدراسة بخصوص نقلها للفايروسات باستثناء الجنس Phylloxera المتغذية أفراده على الأعناب، فيما تضم العائلة Adelgidae ما يقرب من 45 نوعا تتغذى على الصنوبريات ولم يسجل وجود أية ناقلات للفايروسات فيها. تبرز أهمية حشرات المن في نقل الفايروسات بانه من بين 600 نوع فايروسي منقول بناقلات لافقارية فأن ما يقرب من %50 منها تنقل بحشرات المن، (الجدول 1).
ويتم المن دورة حياته بالتبادل بين عائل أساسي وآخر ثانوي ويمر بأشكال حيوية متعددة خلال الدورة تتباين حسب نوع المن وطبيعة المناخ السائد فقد يشتي بشكل بيض أو بأشكال ولودة عذرية Parthenopenetic viviparous، وتقضي بعض أنواع المن دورة حياتها على نوع نباتي واحد أو على عدة أنواع تتبع جنسا نباتيا واحدا، ويعد من الخوخ الأخضر Myzus persicae أحد أخطر أنواع المن الناقلة لفايروسات النبات وتعود النباتات الأساسية العائلة له إلى جنس Prunus الذي يضم أنواع اللوزيات إلا أن مداه العائلي يمتد ليشمل أنواع نباتية تعود إلى ما يزيد عن 50 عائلة نباتية.
يمتلك المن أجزاء فم مكونة من الرمح المرن Stylet والذي يتكون من زوج من الفكوك السفلى الخارجية Mandible وزوج من الفكوك المساعدة الداخلية Maxillae ويحمل الرمح داخل تجويف موجود في الشفة السفلى Labium ويبرز فقط أثناء التغذية وتوجد عليه قرب نهايته البعيدة بروزات حادة شبيهة بالأسنان، (الشكل 3) وعند انطباق الفكوك السفلى الخارجية مع الفكوك المساعدة الداخلية تتكون قناتين هما قناة اللعاب Salivary canal وقناة الغذاء Food Canal حيث يتم قذف اللعاب وحقنه في النبات عن طريق القناة اللعابية ومن ثم سحب الغذاء عن طريق قناة الغذاء، (الشكل 4).
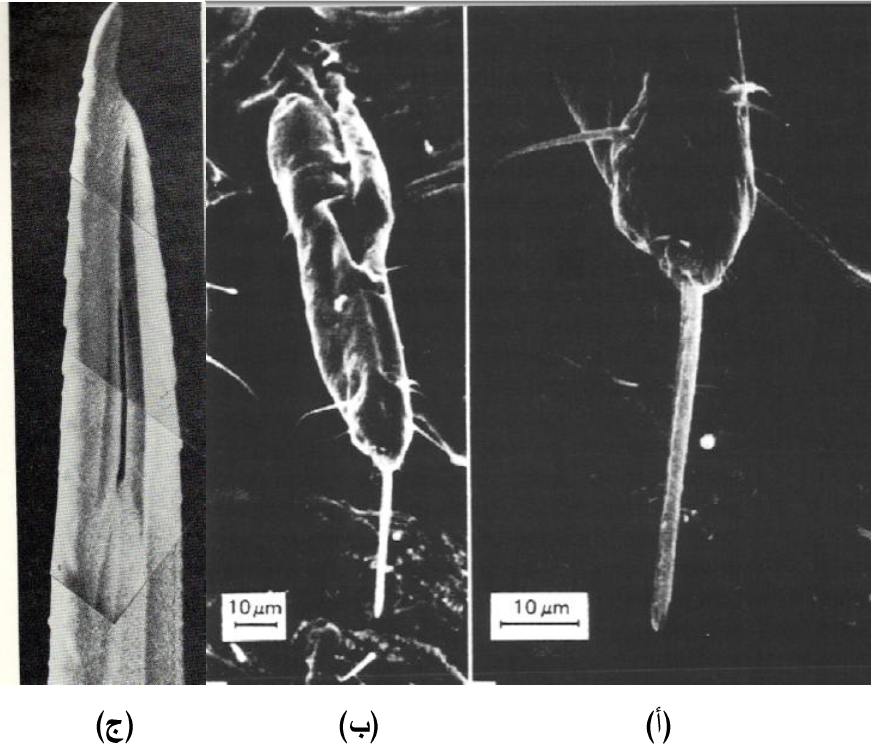
الشكل (3): مظهر خارجي لرمح حشرة من الخوخ الاخضر مصور بالمجهر الالكتروني بتجميد الحشرة في النتروجين المسال بعد سحب رمحها مباشرة من النبات. (أ) الشفة السفلى وقد برز منها الرمح وتلاحظ الشعيرات الحسية الخشنة على سطحها (ب) الرمح بارزا من الشفة السفلى ومغروزا في النبات للتغذية ويلاحظ عدم دخول الشفة السفلى في النبات (ج) قمة الرمح وتلاحظ عليها الاضلاع المسننة وتراكب اجزاء الرمح تماما في النهاية البعيدة بانطباق الفكوك الخارجية والمساعدة.
الشكل مقتبس من Hull (2002) و Gibbs و Harrison (1980).

الشكل (4): مقطع عرضي لرمح حشرة المن Hyperomyzus factucae مصور بالمجهر الالكتروني يبين قناة الغذاء وقناة اللعاب المتكونتين بانغلاق الفكوك المساعدة الداخلية مع وجود قناتي أعصاب في الفكين الخارجيين.
الشكل مقتبس من Gibbs و Harrison (1980).
أ- سلوك التغذية عند المن
بعد استقرار الحشرة على سطح الورقة واتخاذها لوضع التغذية تبدأ بإفراز قطرات من لعاب هلامي القوام ثم يخترق الرمح طبقة البشرة بإجراء طعنات استكشافية Exploratory probes وقد يتغذى المن خلالها مؤقتا ثم يستمر الاختراق إلى الطبقات النباتية الأعمق مع تكوين غشاء لعابي هلامي حول الرمح وعادة ما يدخل الرمح بارزا من الشفة السفلى ملتويا بين الخلايا من دون اختراقها ليصل إلى أنابيب اللحاء وقد يستغرق ذلك دقائق أو ساعات، ويسبب الضغط الذي يسلطه الجدار الخلوي للخلية وهو يقاوم دخول الرمح انكشاف نهاية قمته أي نهايتي قناتي اللعاب والغذاء، (الشكل 5).
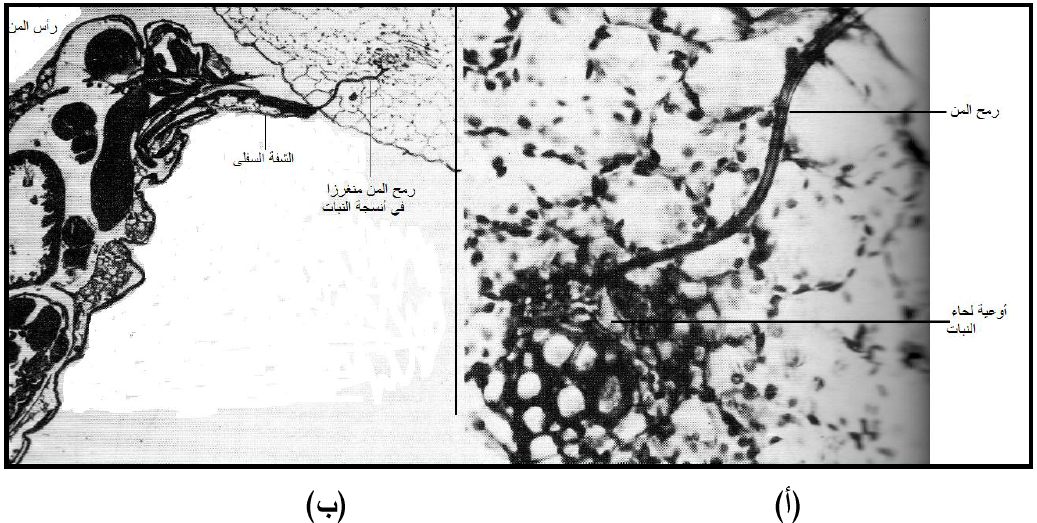
الشكل (5): مقطع في نسيج نباتي أثناء تغذية المن حيث يظهر الرمح مخترقا للأنسجة ليصل إلى اللحاء (أ) انغراز الرمح بين الخلايا بشكل ملتوي باحثا عن اللحاء (ب) رأس حشرة المن والرمح منغرز في النسيج النباتي بارزا من الشفة السفلى التي لا تدخل النبات.
الشكل مقتبس من Gibbs و (1980) Harrison.
نجح الباحثون في إجراء مراقبة الكترونية بالتيارين الترددي والمستمر للمن خلال تغذيته على اللحاء وتم الحصول على معلومات هامة عن سلوكه التغذوي ومواقع تغذيته وذلك باستعمال نوعين من نظم المراقبة وتبين قيام المن بإفراز اللعاب المائي القوام أثناء تغذيته على اللحاء وهذا اللعاب هو الذي يوصل الفايروسات الدوارة والمتكاثرة إلى اللحاء ويساعد أيضا في سحبها منه ولو كان اللعاب هلاميا لاستحالت هذه العملية، أما اللعاب الهلامي فأن المن يفرزه بعد سحب الرمح من النسيج النباتي عند انتهاء التغذية والذي يعمل على إغلاق الجرح الذي تكون نتيجة غرز الرمح في النسيج النباتي وهي عملية إصلاح للضرر الذي تسببه تغذية المن في أنابيب اللحاء والخلايا المحيطة حول مسار الرمح، يبين الشكل (6) مخططا للغشاء اللعابي Stylet Sheath المتكون من اللعاب الهلامي الذي يفرزه المن حول مسار الرمح بعد انتهاء التغذية وبذلك يتضح أن اللعاب المفرز في اللحاء هو مائي القوام يمكن للحشرة حقنه وإعادة ارتشافه، ووجد أن حشرة من الخوخ الأخضر تتمكن من سحب ما يقرب من 10 ميكروليتر/ساعة من عصير النبات خلال الساعة الأولى من التغذية لتزداد إلى 40 ميكروليتر/ساعة عند استمرار التغذية، وهذا يعني نظريا أن كل حشرة تسحب لترا واحدا من عصير النبات إذا تغذت لمدة 25000 ساعة أو أن كل 25000 حشرة تسحب خلال ساعة من التغذية لترا من العصير النباتي.
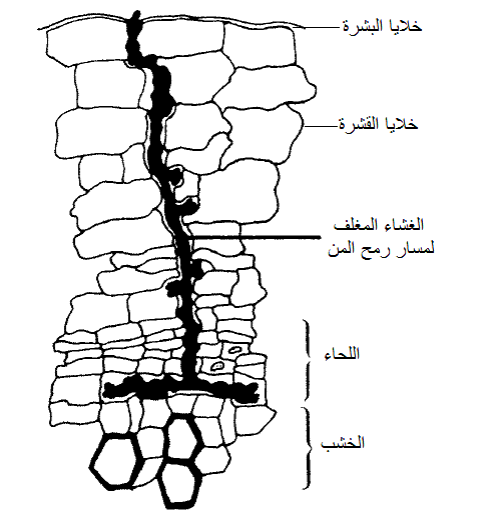
الشكل (6): الغشاء المغلف لمسار رمح المن والمكون من اللعاب الهلامي بعد انتهاء التغذية على اللحاء.
الشكل مقتبس من Hull 2002)).
ب- تأثير النبات العائل على نقل الفايروسات بحشرات المن
تؤثر خواص النباتات على السلوك التغذوي للمن وبالتالي على نقله للفايروسات حيث وجد أن المقاومة التي تبديها بعض النباتات الصليبية ضد حشرة من الصليبيات Brevicoryne brassicae تعتمد على طبيعة الشمع المغلف لسطح الأوراق، كذلك تؤثر كثافة الشعيرات النباتية Trichomes التي تغطي أوراق فول الصويا على السلوك المسنبري Probing behavior للعديد من أنواع المن وعليه فأن انتشار فايروس موزائيك فول الصويا (SMV) في الحقول يتناسب عكسيا مع كثافة تلك الشعيرات، كذلك تؤثر المواد الكيميائية الموجودة في بعض النباتات على النقل بسبب جذبها أو طردها أو تثبيطها لتغذية المن فمثلا تحفز مادة "السنجرين" Syngrin وهي كلايكوسيد زيت الخردل Mustard oil glycoside والموجودة طبيعيا في أوراق النباتات الصليبية تغذية المن المتخصص على الصليبيات ولكنها تثبط تغذية أنواع أخرى غير متخصصة على هذه النباتات، كذلك فأن إصابة بعض أنواع النباتات بالفايروسات تجعلها أكثر ملائمة لتغذية وتكاثر حشرات المن فعند تغذية "من الباقلاء الأسود" على البنجر السكري المصاب بفايروس موزائيك البنجر (BMV) فأنه ينتج المزيد من الأمهات الجديدة مقارنة بأعدادها عند تغذيته على النباتات السليمة وبذلك يحصل ازدحام شديد Overcrowding للحشرات بوقت مبكر على النباتات المصابة مما يجعلها تبدأ الهجرة مبكرا مقارنة بالنباتات السليمة مما يؤدي إلى نشر الفايروس، ويفضل من الخوخ الأخضر نباتات البنجر المصابة بفايروس اصفرار البنجر (BYV) في التغذية والتكاثر كما أنه يعيش عليها فترة حياة أطول مقارنة بالمن المتغذي على النباتات السليمة، كما وجد أن حشرات المن المجنحة المتغذية على الشوفان المصاب بفايروس التقزم الاصفر للشعير (BYDV) تحسن نضجها بنسبة 85% فيما قصرت فترة حياة المن المتغذي على الحنطة المصابة بنفس الفايروسي المذكور.
ج- تأثير الظروف البيئية على نقل الفايروسات بحشرات المن
تلعب الظروف البيئية وخاصة الحرارة والرطوبة والضوء والرياح دورا مهما في تغذية وحركة المن وبالتالي على كفاءته في نقل الفايروسات، فوجد مثلا زيادة كفاءة نقل فايروس تقزم يوسفي الساتسوما (SDV) بالمن عند 20-22م مقارنة بدرجات 10-11م أو فوق 29م، وتزيد الرطوبة العالية من كفاءة نقل فايروسي البطاطا واي (PVY) والتفاف أوراق البطاطا (PLRV) بالعديد من أنواع المن المتغذية على البطاطا، كما تؤثر نوعية الضوء على حركة نوعين من فايروسات Luteoviruses المتطفلة على الطماطة وبالتالي على اكتسابهما من قبل من الخوخ الأخضر وتؤثر الظروف البيئية أيضا وبشكل غير مباشر على كفاءة نقل الفايروسات بالمن من خلال تأثيرها على تقبلية النبات للفايروسات وتركيزها فيه.
د- العلاقات بين حشرات المن وفايروسات النبات
تعد العلاقة بين المن والفايروسات هي أعقد أنواع العلاقات بين الحشرات وفايروسات النبات حيث سجلت أربعة أنواع من العلاقات التخصصية بينهما وهي:
1- العلاقة غير الباقية
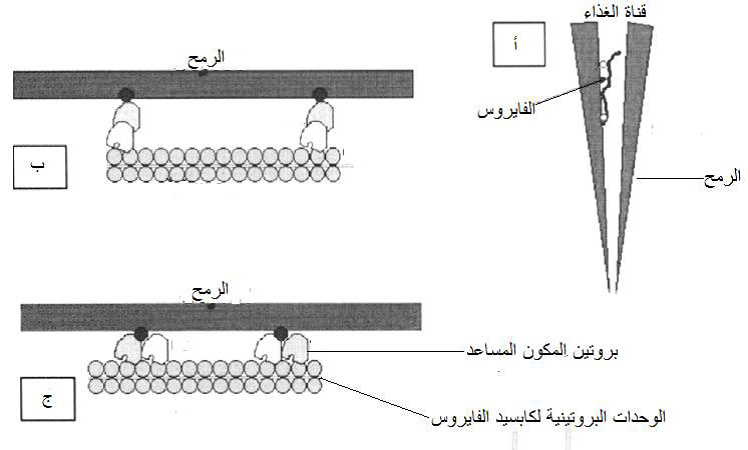
تظهر العلاقة غير الباقية Non – persistent relationship في الفايروسات غير الباقية Non-persistent viruses وهي الفايروسات المحمولة خارجيا بالناقل Externally-borne viruses أي الفايروسات المحمولة على الرمح، وتسمى هذه العلاقة أيضا "غير الدوارة Non- circulative relationship وتعد إحدى أهم العلاقات القائمة بين المن والفايروسات والتي تساهم بشكل كبير في نقل الفايروسات نقلا أفقيا في الحقول، حيث أنه من بين 290 نوعا فايروسيا منقولا بالمن فأن القسم الأكبر منها منقول بهذه الطريقة وتنتمي الأنواع الفايروسية المنقولة بهذه الطريقة إلى خمسة أجناس هي Alfamovirus وPotyvirus, Macluravirus, Fabavirus, Cucumovirus والتي تضم مختلف أشكال الفايروسات ففيها العصوي والايزومتري وكذلك كل أنواع الأحماض النووية الفايروسية مفردة أو مزدوجة الخيط. ولنجاح هذا النقل فأنه لا يتطلب اختراق رمح المن لأكثر من طبقة خلايا بشرة النبات المصاب لاكتساب الفايروس ويحصل ذلك نتيجة سلوك التغذية القصيرة الاستكشافية Expletory probing feed التي يمارسها المن في بداية وصوله إلى سطح النبات حيث يلتقط الفايروس وترتبط جسيماته بالبطانة الكيوتكلية للجزء الداخلي من القمة الأمامية للرمح Stylet tip وقد اثبت ذلك عمليا حيث أدت معاملة الرمح بالفورمالين أو بالأشعة فوق البنفسجية إلى إبطال هذا النقل نتيجة إتلاف الفايروس مما دل على وجوده في هذه المنطقة. وضعت فرضيتان تفسر ان الية اكتساب هذه الفايروسات وكيفية حملها في الرمح وهما (1) الية قذف الفريونات Egestion من القناة الغذائية للرمح وإرجاعها مع الغذاء إلى مقدمة الرمح بحركة انعكاسية للمضخة الساحبة Sucking pump للحشرة (2) الية إفراز اللعاب Salvation وحقنه في النبات من قبل الحشرة، وبذلك يتم الاحتفاظ بهذه الفايروسات في مفصل قناتي الغذاء والعاب أي في الفسحة المشتركة بينهما عند قمة الرمح، (الشكل 7) حيث توفرت الكثير من الأدلة التي تدعم ذلك إذ أظهر المجهر الالكتروني وجود جسيمات فايروسي نقش التبغ (TEV) والتبرقش العرقي للتبغ (TVMV) مرتبطة بالكيوتكل الخارجي Epicuticle المبطن لقناة الغذاء فيما كانت هناك أعدادا قليلة من الجسيمات مرتبطة بمنطقتي التجويف الفمي Cibarium ومقدمته Precibarium، وأظهرت تقانات التعليم الشعاعي باستعمال جسيمات فايروسية معلمة اشعاعيا Radioactively labeled viruses أن هذه الجسيمات ظهرت على الأسطح الداخلية للثلث الطرفي لقناة الغذاء في نهاية الرمح وأنه يتم إطلاقها وحقنها في النبات عند إفراز العاب وضخه في النبات.
يحدث الارتباط بين الفايروس والسطح الداخلي لرمح الحشرة بطريقتين (1) الارتباط المباشر بين الكابسيد والكيوتكل المبطن للرمح حيث يعمل جزء معين من الكابسيد عمل الرابط بين جسيمة الفايروس والكيونكل وأثبت ذلك مع فايروسي موزائيك الجت (AMV) وموزائيك الخيار (CMV) بإمكانية نقل الفايروسين عند تغذية المن على المحلول الفايروسي النقي لكل منهما عبر غشاء من دون إضافة أية بروتينات أو مواد أخرى، وتم تأكيد ذلك أيضا عند إعادة تركيب جينوم فايروس موزائيك التبغ (TMV) غير المنقول بالمن مع كابسيد فايروس اسبيرمي الطماطة (TAV) المنقول بالمن حيث أدى ذلك إلى نجاح نقل الفايروس الأول المهجن. وجد أن لبعض الأحماض الامينية ولمواقعها في الكابسيد دورا في تخصصية الربط ولوحظ ذلك في العلاقة بين حشرة "من القطن"A. gossypii وفايروس موزائيك الخيار إذ وجد أن مواقع الأحماض الامينية 25 و 129 و168 و 162 و 214 وفق تسلسلها في الكابسيد هي المواقع الحاكمة لنقل الفايروس المذكور بواسطة من الخوخ الأخضر ويفسر دورها التخصصي بمسئوليتها عن تحديد طبيعة التفاف ببتيدات الغطاء البروتيني أو في ترسيخ الثباتية الفيزيائية للفايروس داخل جسم المن الناقل (2) الارتباط غير المباشر للجسيمة الفايروسية وكيوتكل الرمح عبر وسيط هو البروتين المساعد Helper protien الفايروسي المتخصص بربط كابسيد الفايروس مع رمح المن والذي يشكل جسرا رابطا بينهما وهو المسئول عن النقل التخصصي للفايروسات بالمن ويوجد قريبا من النهاية النتروجينية للكابسيد حيث يضم مجموعة مكونة من ثلاث أحماض أمينية هي الأسبرجين والألانين والكلايسين ومجموعة رباعية تتكون من الأحماض الامينية السستين والثريونين والأيزوليوسين واللايسين وهما مجموعتان ضروريتان للنقل، ويطلق على هذا البروتين أيضا مصطلح "المكون المساعد "Helper protien أو "العامل المساعد" Helper factor أو يسمى "عامل النقل بحشرات المن Aphid transmission factor وهو بروتين وظيفي يشفره الفايروس وتخلقه الخلية المصابة، (الشكلين 8 و 9) وقد سجل وجود هذا البروتين في فيروسات "البوتي" Polyviruses والذي يتباين في حجمه ووزنه الجزيئي حسب نوع الفايروس فهو في فايروس التبرقش العرقي للتبغ (TVMV) بوزن جزيئي 53 كيلو دالتون (KDa) وفي فايروس البطاطا واي (PVY) بوزن جزيئي 58 كيلو دالتون. إن البروتين المساعد هو أصلا جزء بروتيني ينشطر من البروتين الكبير Polyprotein الذي تشفره فايروسات Potyviruses والذي يطلق عليه اختصارا HC-Pro هو رمز مشتق من مصطلح" المكون المساعد – إنزيم تحليل البروتين" Helper Component-Protease وهذا يعني أنه يتكون من جزأين الأول هو البروتين المساعد للنقل والثاني هو الأنزيم المحلل للبروتين لذا فهو ذو وظيفتين حيويتين ويرتبط في فايروسات "البوتي" ارتباطا غير تخصصي بالحامض النووي الفايروسي من خلال منطقتي ربط بالحامض، وأظهرت الدراسات أنه يفضل الارتباط مع الرنا أكثر من ميله للارتباط بالدنا الفايروسي، كما وجد أنه يساعد أيضا في نقل أنواعا أخرى من الفايروسات غير المنقولة بالمن عند وجود أحد فايروسات "البوتي" في إصابة خليطة مع فايروسات أخرى لا تنقل بالمن في نفس النبات حيث يكتسب المن ذلك البروتين إما خلال تغذية الاكتساب لفايروس "البوتي" أو قبلها مباشرة ليصبح قادرا على نقل الفايروس الأخر الذي لم يكن أصلا منقولا بالمن، أما إذا اكتسبه المن بعد اكتساب فايروس "البوتي" عندها يفشل نقل الفايروس الأخر، وهذه الظاهرة هي التي نبهت الباحثين على وجود هذا البروتين، ويسمى فايروس "البوتي" الذي يساعد فايروسا آخر لنقله بحشرات المن "الفايروس المساعد" Helper Virus وهي ظاهرة شائعة في هذه الفايروسات إلا أنها متخصصة في دورها النقلي بمعنى أنه ليس كل فايروسات البوتي ستساعد أي فايروس من مجموعة أخرى للنقل بالمن إن وجدا معا في إصابة خليطة، وتوجد العديد من الأمثلة على ذلك ومنها دور فايروس البطاطا واي (PVY) في نقل فايروس موزائيك أوكيوبا البطاطا (PAMV). أمكن تنقية وتشخيص البروتينات المساعدة لبعض فايروسات البوتي ومنها فايروسات التبقع الحلقي للبابايا (PRSV) وموزائيك الرقي الثاني (2-WMV) و الموزائيك الاصفر لقرع الزكيني (ZYMV) وموزائيك الشلغم (TuMV).
أما بخصوص آلية إطلاق الفايروسات غير الباقية من الرمح بعد اكتسابها فقد وضعت فرضيتين لتفسير الكيفية التي تتحرر بها هذه الفايروسات من الرمح وهما (1) الية الابتلاع والحقن Injestion-egestion mechanism وهي التي تفترض إطلاق الجسيمات الفايروسية من الرمح نتيجة عمليتي الاسترجاع Regugitation وإفراز العاب Salivation التي تقوم بهما الحشرة (2) إطلاق الجسيمات بتأثير قذف العاب فقط ويساعد على ذلك أن قناتي العاب والغذاء متحدتان قرب قمة الرمح الطرفية. ويلخص الجدول (2) مواصفات النقل غير الباقي بحشرات المن.
الجدول (2): العلاقات الحيوية بين فايروسات النبات وحشرات المن

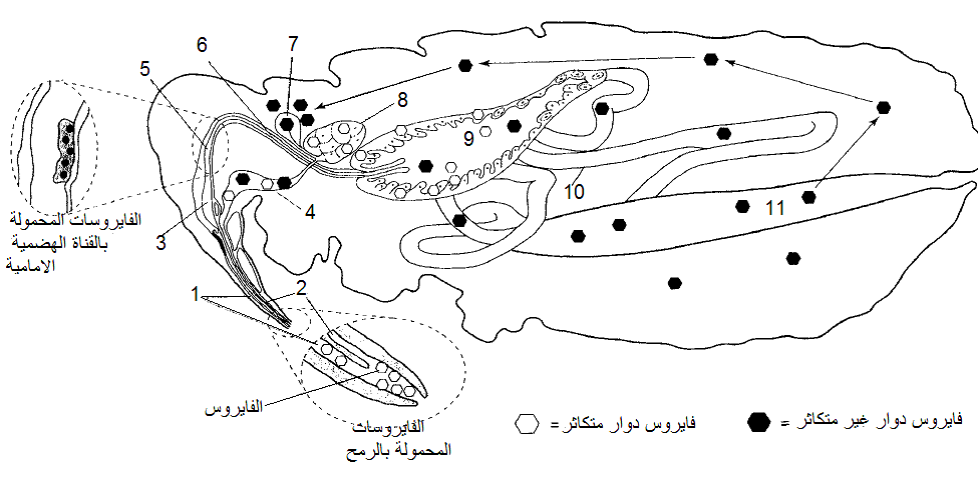
الشكل (7): اليات نقل فايروسات النبات بواسطة الحشرات الثاقبة الماصة.
الشكل مقتبس من Hull 2002.
يبين الشكل تشريح القناة الهضمية والجهاز اللعابي والمواقع الاخرى ذات الصلة بنقل الفايروسات. تظهر الصورة المكبرة السفلى توضيحا للنهاية البعيدة للرمح حيث تنتهي قناتي الغذاء (1) واللعاب (2) في فجوة مشتركة في نهاية الرمح. إن الألية المقترحة لنقل الفايروسات غير الباقية المحمولة بالرمح هي الاحتفاظ بها في القمة البعيدة للرمح ثم يتم إطلاقها بواسطة العاب المفرز عندما تحقنه الحشرة في النبات خلال التغذية. تظهر الصورة المكبرة العليا (إلى اليسار) موقع وجود الفايروسات شبه الباقية المحمولة بالقناة الهضمية الأمامية Foregut-borne viruses حيث ترتبط الجسيمات الفايروسية بالكيوتكل المبطن للقناة منطمرة في مادة مالئة تسمى Matrix material مرتبطة بالكيوتكلل لا يعرف تركيبها الدقيق، وتوجد في نهاية هذه القناة المضخة الماصة (3) والبلعوم (5) والمريء (6). أما الفايروسات الدوارة غير المتكاثرة فأنها ستعبر القناة الهضمية الأمامية إلى الجزء الأمامي من القناة الوسطى (9) ثم إلى الجزء الخلفي للقناة الوسطى (10) ثم إلى القناة الهضمية الخلفية (11)، وهي لا تصيب خلايا القناة الهضمية ولكنها تعبر خلال خلايا الجزء الخلفي للقناة الوسطى والأمامي للقناة الخلفية وتتحرر إلى الدم (الهيموسيل) Hemocoel ثم تصل إلى الغدد اللعابية المساعدة (7) وتنتقل عبر خلاياها ثم تتحرر إلى القناة اللعابية (2). أما الفايروسات المتكاثرة فهي تصيب خلايا القناة الهضمية الوسطى ثم تصيب أنسجة أخرى لاحقة لترتبط بعدها بالغدد اللعابية الأساسية (8) وربما بالغدد اللعابية المساعدة قبل أن تتحرر في القناة العابية نتيجة ضخها بواسطة مضخة اللعاب (4) كما هو الحال مع الفايروسات الدوارة غير المتكاثرة.
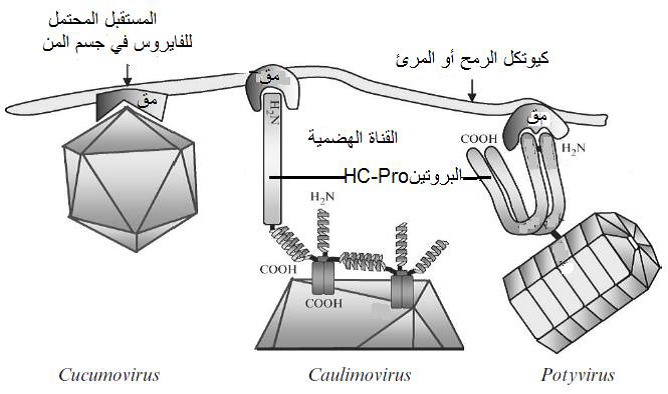
الشكل (8): نموذج افتراضي لألية ارتباط فايروسات اجناس Potyvirus و Caulimovirus و Cucumovirus غير الباقية وشبه الباقية المنقولة بالمن على كيوتكل الرمح او المريء. يلاحظ الاحتفاظ بالفايروسات بواسطة مستقبل واحد او اكثر موجود على الكيوتكل يسمى مستقبل المن Aphid receptor والذي اعطى الرمز "مق" حيث تحتاج فايروسات الجنسين Caulimovirus و Potyvirus الى البروتين الفايروسي المتخصص بالنقل HC-Pro لكي تدمص على كيوتكل الرمح او المريء، اما مع فايروسات الجنس Cucumovirus فيكون الارتباط مباشر ومن دون تدخل لاي بروتين فايروسي.
الشكل مقتبس من Foster واخرون (2008).
الشكل (9): الأليات المحتملة لربط بروتين المكون المساعد الذي يشفره الفايروس والمسمى البروتين HC-Pro" مع كابسيد فايروسات البوتي Potyviruses ومع رمح المن (أ) موقع الجسيمات الفايروسية داخل الجزء الأمامي من الرمح وتحديدا بداية قناة الغذاء (ب) نموذج يفترض الارتباط بين جزئ البروتين حيث يرتبط الجزء الأول مع مستقبل موجود على الرمح فيما يرتبط الثاني مع وحدة بروتينية من وحدات الكابسيد (ج) نموذج يفترض الحاجة إلى "مثنى" Dimer للربط مع المستقبل الموجود على الرمح إذ يرتبط كلا جزئي HC-Pro مع الوحدات البروتينية للكابسيد.
الشكل مقتبس من Hull (2002).
2- العلاقة شبه الباقية
العلاقة شبه الباقية Semi-persistent relationship هي الأقل شيوعا من بقية العلاقات القائمة مع المن وهي تجمع في صفاتها بين العلاقة غير الباقية والعلاقة الباقية الدوارة وكما مبين في الجدول (2) وسجلت هذه العلاقة مع خمسة أجناس فايروسية هي Caulimovirus و Closterovirus و Sequivirus و Carlavirus و Vitivirus الا ان أكثرها دراسة هي الأنواع التابعة للجنسين الأول والثاني وخاصة فايروسي اصفرار البنجر (BYV) وترستيزا الحمضيات (CTV) المنتميان للجنس الثاني ويصيبان اللحاء، أما الفايروسات التابعة للجنس الأول ومنها فايروس موزائيك القرنابيط (CaMV) فهي موجودة في معظم أنسجة النبات وعند نقلها بهذه الطريقة فأنها تحتاج إلى وجود البروتين المساعد والذي يتدخل في نقل فايروسات الجنس Sequivirus إلا أنه لم يتم تشخيصه لحد الآن. لا تظهر الفايروسات المنقولة بهذه الطريقة صفات نقل متجانسة بينها لذا فمن الصعب وضع مواصفات محددة لهذه العلاقة ولكنها عموما ذات فترة تغذية اكتساب وتلقيح طويلة تستغرق عدة ساعات وكلما ازدادت تزداد كفاءة النقل وذلك لان معظم الفايروسات شبه الباقية توجد في اللحاء مما يتطلب وصول الرمح إليه، ولا توجد فترة حضانة لان مواقع الاحتفاظ بهذه الفايروسات في جسم المن تكون في بداية القناة الهضمية الأمامية، (الشكل 7) وهناك أدلة على أن الفايروسات شبه الباقية تحتاج إلى البروتين المساعد لنقلها التخصصي بحشرات المن إلا أن تركيب هذا البروتين يختلف عن تركيب مثيله في الفايروسات غير الباقية فهو أكثر تعقيدا حيث وجد أن البروتين المساعد الذي يحتاجه فايروس موزائيك القرنابيط لنقله بحشرة من الخوخ الأخضر يتكون أصلا من نوعين من بروتين الغطاء الفايروسي الأول بحجم 18 كيلو دالتون وهو ناتج المنطقة الجينومية الثانية ORFll والثاني بحجم 15 كيلو دالتون وهو ناتج المنطقة الجينومية الثالثة ORFlll وهو يجب أن يكتسب من قبل المن قبل أو خلال التغذية كي ينقل الفايروس، ووجد أن لبعض البروتينات الاخرى التي تنتجها فايروسات الجنس Caulimovirus قدرة الارتباط التكاملي مع هذا البروتين ومساعدته في مهمة النقل، (الشكل 8) ولوحظت في هذه العلاقة أيضا ظاهرة النقل المساعد التي وجدت في الفايروسات غير الباقية حيث أن فايروس الترقط الاصفر للجزر الأبيض (PYFV) لا ينقل بالمن ولكن عند وجوده في النبات بإصابة خليطة مع فايروس اصفرار البقدونس الافرنجي (AYV) المنقول بالمن عندها يصبح المن الناقل Cavariella aegopodii ناقلا له حيث يعمل الأخير فايروسا مساعدا للأول وهذا يشير إلى تخصصية هذا النقل حيث يمتلك الفايروس الثاني موقع ارتباطي تخصصي في المعي الأمامي للحشرة، وهناك أدلة على أن فايروسات Carlaviruses وClosteroviruses تحتاج إلى بروتينات مساعدة لنقلها بالمن وسجل ذلك مع فايروس الهرقلية الكامن HLV (الهرقلية نوع من الازهار من العائلة الخيمية) وهو عصوي مرن بطول 730 نانومتر حيث يعتمد في نقله على فايروس الهرقلية السادس (HV-6) العصوي بطول 1600 نانومتر وكلاهما ينتميان لجنس Closterovirus إذ يرتبطان معا من نهايتيهما طرف لطرف عند نقلهما بالحشرة وهي حالة غريبة من الارتباط لم تسجل مع فايروسات أخرى، ويبدو أن الجزء البروتيني الذي يغطي النهاية 5 لجينوم فايروسات Closteroviruses هو المسئول عن نقلها تخصصيا بالمن.
تظهر في هذه العلاقة ظاهرة "النقل المزدوج" Bimodal transmission والذي يتم فيه نقل فايروس نباتي معين بالطريقتين غير الباقية وشبه الباقية بنفس نوع المن، ولكن رغم تسجيل هذه الظاهرة مع عدد قليل من الفايروسات إلا أن بعض الباحثين يشكك في وجودها معللين بأنها قد تظهر نتيجة لاختلاف في العوامل المحيطة بعملية النقل وخاصة التفاعل بين المن الناقل والنبات العائل للفايروس.
3- العلاقة الباقية
العلاقة الباقية Persistent relationship والتي تسمى أيضا " العلاقة الدوارة" Circulative relationship هي من العلاقات المهمة بين العديد من أنواع الفايروسات وحشرات المن ويطلق على الفايروسات التي تمتاز بهذه العلاقة "الفايروسات الدوارة الباقية" Persistent circulative viruses وتسمى أيضا "الفايروسات المحمولة داخليا بالناقل" Internally-borne viruses أي داخل القناة الهضمية وتجويف الجسم والدم، (الشكل 7) وتنقل بهذه الطريقة الأنواع التابعة لأجناس Nanovirus، عائلة Circovridaeو Luteovirus و Pollerovirus و Enarmovirus، عائلة Luteoviridae ،وتلعب هذه العلاقة دورا مهما في النقل الأفقي لعدد كبير من أنواع الفايروسات الخطرة على الإنتاج الزراعي والتي تسبب أعراض الاصفرار والتفاف الأوراق غالبا لأنها تتطفل على اللحاء كما أنها مسئولة عن النقل القريب والبعيد للفايروسات، ويلخص الجدول (2) المواصفات الرئيسية لهذه العلاقة. يزداد مستوى التخصصية في هذه العلاقة بين الفايروس والمن مقارنة بالعلاقات السابقة حيث لا ينقل النوع الفايروسي الباقي إلا بنوع معين من المن أو ربما بعدة أنواع قليلة مما يشير إلى التخصصية العالية فيها، بينما ينقل الفايروس غير الباقي بعدد كبير من أنواع المن.
يتم في هذه العلاقة ابتلاع الجسيمات الفايروسية من قبل الحشرة عند التغذية الطويلة لتنقل مع العصير النباتي إلى الأمعاء ثم تعبر إلى الدم (هيمولمف الحشرة) وتحديدا من المعي الخلفي للحشرة ثم تصل إلى الغدد اللعابية لتحقن مع العاب في النبات الجديد الذي تتغذى عليه الحشرة، لذا فإن دورتها هذه في جسم الحشرة يتطلب أن تعبر حاجزين هما جدار الأمعاء وجدر الغدد اللعابية وفترة حضانة قد تستغرق 12 ساعة- عدة أيام حسب نوع الفايروس، (الشكل 10) كما أن المدة المعتادة لاكتساب هذه الفايروسات هي عدة ساعات رغم تسجيل فترة اكتساب مع بعض الفايروسات لا تستغرق أكثر من خمسة دقائق في حدها الأدنى، وتحتاج الحشرة لفترة تغذية لمدة 10-30 دقيقة لتلقيح النبات السليم، وتبقى الحشرة الحاملة للفايروس قادرة على نقله لعدة أيام. ولفهم كيفية عبور هذه الفايروسات للأغشية الداخلية للحشرة فقد درست آلية عبور جسيمات بعض أنواع فايروسات Luteoviruses لأغشية أنسجة حشرات المن ومنها فايروس التقزم الاصفر للحبوب (CYDV-RPV) والذي كان
يسمى سابقا فايروس التقزم الاصفر للشعير (BYDV- RPV) حيث ترتبط جسيماته بالأغشية الخلوية لخلايا المعي الخلفي للمنRopalosiphon padi ومن المحتمل أن تدخل إليها بواسطة عملية الابتلاع Endocytosis حيث تتجمع الجسيمات الفايروسية أولا داخل نقر أو حويصلات مغلفة Coated pits or vesicles ثم تتراكم في حويصلات أنبوبية وفي اللايسوسومات Lysosomes لتطلق إلى الهيموسيل عن طريق اتحاد الحويصلات الأنبوبية مع الغشاء البلازمي القاعدي Basal plasmalemma، ودرست آلية عبور فايروسي اصفرار البنجر الغربي (BWYV) والتفاف أوراق البطاطا(PLRV) من الدم إلى الأغشية اللعابية لحشرة من الخوخ الأخضر ووجدت جسيماتهما في فجوات الغشاء البلازمي Plasmalemma invaginations لخلايا الغدد اللعابية المساعدة وكذلك في الحويصلات الأنبوبية في سايتوبلازم الخلايا المجاورة للغدد اللعابية وفي النقر المغطاة المرتبطة بالغشاء المسمى بالغشاء القناتي Canal membrane حيث يتم عبور هذه الأغشية بواسطة مستقبل بروتيني "بالية الابتلاع الميسرة بالمستقبل"Receptor mediated endocytosis وتوفرت أدلة على وجود البروتين Read through protein وهو جزء من كابسيد هذين الفايروسين والذي يعد ضروريا لنقلهما بالمن حيث أن الفايروسات التي لا تمتلكه يمكنها أن تصل إلى الهيموسيل بعد عبورها لجدار الأمعاء ولكنها تفشل في عبور جدر الغدد اللعابية، كما اكتشفت أنواع أخرى من البروتينات التي تلعب دورا في ربط الجسيمات الفايروسية بأغشية الحشرات فقد عزلت بروتينات من حشرات المن بوزن جزيئي 31 و85 كيلو دالتون حاملة لبعض أنواع فايروسات Luteoviruses، كذلك سجل وجود بروتين السمبيونين Symbionin ذو الوزن الجزيئي 60 كيلو دالتون والذي له قدرة الارتباط بأغشية المن وهو يعمل مع أنواع فايروسية عديدة منها الأنواع التي تعود للجنس Luteowus وهو ينتج من قبل البكتريا المتعايشة داخليا في أمعاء المن والتي تعود للجنس Buchnella حيث وجد في هيمولمف المن وهو يتبع مجموعة بروتينات الكابيرون Chaperone المسئولة عن تثبيت هيكلية البروتينات حيث يرتبط مع الجسيمة الفايروسية في منطقة Read through domain الموجودة في الكابسيد، ووجد أنه عند معاملة من الخوخ الأخضر بالمضادات الحيوية أدى ذلك إلى خفض مستويات هذا البروتين في الحشرة وثبط نقلها للفايروس وفقدان تماسك الكابسيد، كما أن لهذا البروتين دورا في ثباتيه الجسيمات الفايروسية داخل هيمولمف الحشرة.
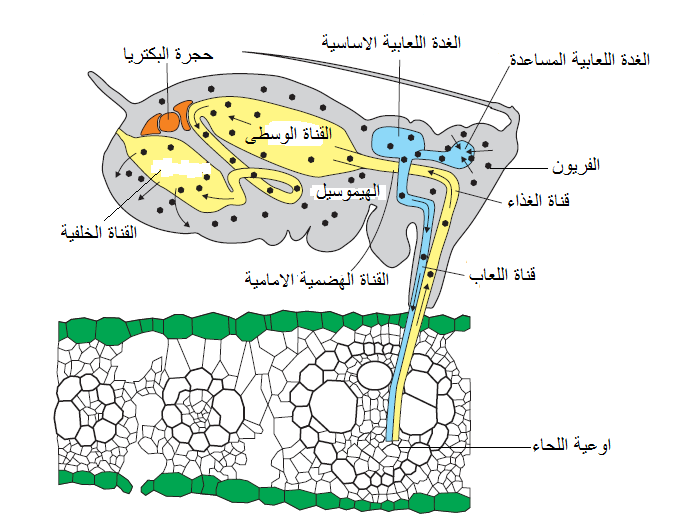
الشكل (10): نموذج للنقل الدوار للفايروسات بواسطة حشرات المن.
تكتسب الحشرة الفريونات من أوعية اللحاء في النبات المصاب وتتجه الفريونات عبر الرمح خلال قناة الغذاء إلى القناة الهضمية الامامية ثم تنقل بالنقل النشط عبر خلايا جدار القناة الخلفية إلى الهيموسيل ثم تنتشر عبر الهيمولمف إلى الغدة اللعابية المساعدة ثم تنقل بالنقل النشط إلى قناة الغدة وتقذف من هناك مع العاب إلى اوعية اللحاء في النبات الجديد.
الشكل مقتبس من Domier 2008))
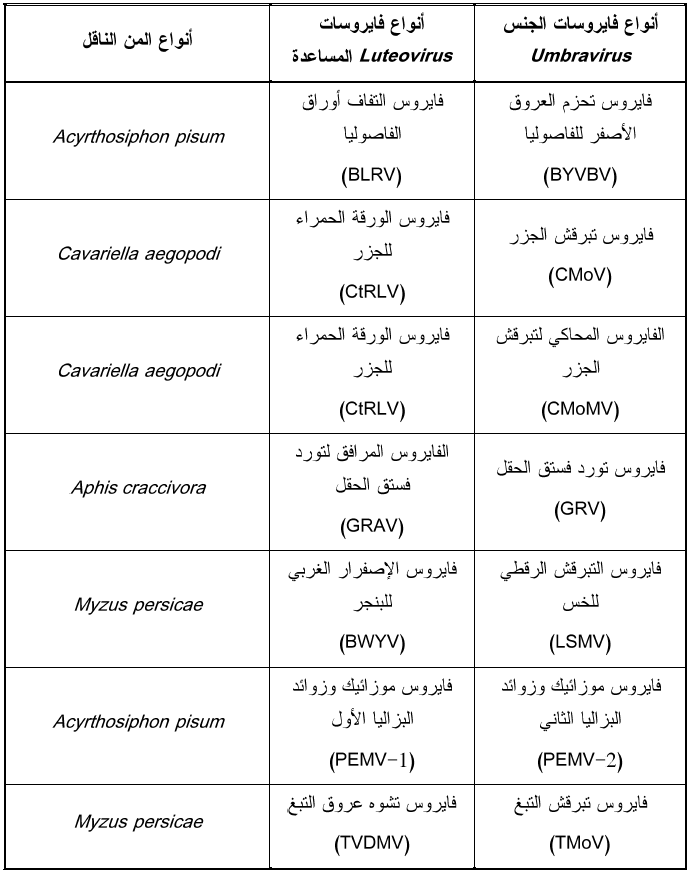
من الظواهر المهمة المرافقة للعلاقة الباقية بين المن والفيروسات هي ظاهرة "النقل الإتكالي" Dependent transmission حيث تحتاج بعض هذه الفايروسات إلى فايروس مساعد أو منشط Helper or Activator Virus موجود مسبقا في النبات كي تنقل بالمن ولوحظت هذه الظاهرة مع فايروس التقزم الاصفر للحبوب (CYDV-RPV) المنتمي للجنس Luteovirus والذي ينقل بكفاءة بواسطة نوع المن R. Padi ولكنه لا ينقل بالمن Sitoboin avenae، أما فايروس اصفرار وتقزم الشعير (BYDV-MAV) فينقل بكفاءة بالأخير ولا ينقل بالأول ولكنهما ينقلان بنفس الكفاءة وبكلا نوعي المن عند وجودهما معا في النبات بإصابة خليطة اذ وجد أن كلا الفايروسين يستطيعان الدخول إلى هيمولمف الناقل غير الكفاء ولكنهما يفشلان في عبور غدده اللعابية ليصلا إلى النبات، ويفسر التأثير المنشط للإصابة الخليطة في نقلهما إلى وجود ما يعرف بالخلط الشكلي Phenotypic mixing وذلك عند تضاعف الفايروسين معا في النبات حيث يتم تغليف بعض جينومات فايروس اصفرار وتقزم الشعير (BYDV-MAV) بالغطاء البروتيني لفايروس التقزم الاصفر للحبوب (CYDV-RPV) وبذلك يسهل نقله بالمن غير الكفء. لوحظت هذه الظاهرة أيضا مع فايروسات Umbraviruses التي لا تمتلك قدرة تشفير غطاؤها البروتيني لذلك ترتبط بفايروس مساعد من فايروسات الجنس Luteovirus ليمنحها الكابسيد الذي يربطها بالمن، ولوحظت أيضا مع فايروسات Nanoviruses التي تحتاج إلى فايروس الاصفرار التماوتي للباقلاء (FBNYV) لنقلها بالمن، كذلك فان فايرويد الدرنة المغزلية للبطاطا (PSTVd) ينقل بالمن عندما تكون نباتات البطاطا مصابة بفايروس التفاف أوراق البطاطا (PLRV) رغم وجود شك بإمكانية تعبئة الفايرويد في كابسيد الفايروس المذكور. تمتاز العلاقة الاتكالية بالمزايا التالية (1) أن كلا الفايروسين ينقلان بالطريقة الباقية غير المتضاعفة (2) ينقل الفايروس الاتكالي ميكانيكيا بينما لا ينقل الفايروس المساعد بهذه الطريقة (3) لا ينقل الفايروس الاتكالي بالمن إلا إذا كان النبات مصاب بكلا الفايروسين إصابة مشتركة حيث يتم تعبئة الفايروس الاتكالي في كابسيد الفايروس المساعد، يبين الجدول (3) سبعة أنواع من الفايروسات الاتكالية وفايروساتها المساعدة.
الجدول (3): الفايروسات الاتكالية التابعة لجنس Umbravirus والفايروسات المساعدة لنقلها بالمن والتابعة للجنس Luteovirus.
4- العلاقة الباقية الدوارة المتضاعفة
العلاقة الباقية الدوارة المتضاعفة Propagative relationship هي العلاقة الأكثر تخصصا بين المن والفايروسات المنقولة بهذه الطريقة والتي تسمى الفايروسات الدوارة المتضاعفة Propagative Viruses وهي الفايروسات التي تتضاعف داخل ناقلاتها الحشرية لذلك فأن هذه العلاقة تحقق النقل الأفقي للفايروسات فضلا عن النقل العمودي لها أيضا عبر أجيال الحشرة، وتمتاز هذه العلاقة بذات المواصفات التي تميز العلاقة الباقية الدوارة والفرق الوحيد بينهما أن الفايروسات في هذه العلاقة تتكاثر في أنسجة ناقلاتها وتحديدا في الدم بينما لا يحصل ذلك في العلاقة الباقية الدوارة، كما تمثل هذه العلاقة حالة نادرة من التطفل المزدوج للفايروس على عائل نباتي وآخر حيواني معا برغم التباين بين الأنسجة الحيوانية والنباتية وهي حالة لم تسجل مع أي من الطفيليات الأخرى. سجلت العديد من الفايروسات المنقولة بهذه الطريقة التابعة لجنسي Cytorhabdovirus وNucleorhabdovirus التابعين لعائلة Rhabdoviridae حيث شوهدت جسيمات فايروسي العرق الاصفر للكسوب (SYVV) والاصفرار التماوتي للخس (LNYV) المنقولين بالمن Hyperomyzus lactucae في نوى وسايتوبلازم خلايا دماغ الحشرة وفي خلايا الغدة تحت المريئيةSubesophageal ganglion وفي خلايا الغدد اللعابية والمبايض والجسم الدهني والعضلات وحجرة الكائنات الدقيقة Mycetome ويبدو أن جسيماتهما تتجمع في نوى الخلايا، وهما ينقلان بواسطة بيض المن المذكور (يستعمل مصطلح Transoviral transmission للإشارة لنقل الفايروسات ببيض الحشرات الحاملة للفايروس) وذلك بعد فترة حضانة طويلة تعتمد على درجات الحرارة، ووجد أن 1% فقط من الحوريات الناتجة من البيض كانت حاملة للفايروس الأول وقادرة على نقله كما تسببت إصابة حشرات المن بالفايروس زيادة في نسب موتها وأمكن نقل الفايروس تسلسليا من حشرة لأخرى بحقن المحلول الفايروسي في الهيمولمف. إن هذه العلاقة هي أقل شيوعا مقارنة بالعلاقات الاخرى ورغم ذلك فهي من العلاقات التي تسبب انتشار ونقل العديد من الفايروسات الخطرة ومنها فايروس التفاف أوراق البطاطا (PLRV) الذي ينقل بالمن بهذه الطريقة حيث وجد أنه عند حقن الحشرات بالمحلول النقي للفايروس المذكور ولمرة واحدة فأن 50% من الحشرات المحقونة أصبحت قادرة على نقله طيلة حياتها بعد فترة حضانة 20 ساعة.
يبدوا مما تقدم أن العلاقات الأربعة مع المن تعتمد على عاملين هما (1) مواقع الاحتفاظ بالفايروس في جسم الحشرة Site of retention (2) مسار الفايروس داخل جسم الحشرة Route of transport. كما تمر دورة نقل الفايروسات بالمن وبغض النظر عن نوع العلاقة بأربع مراحل هي (1) مرحلة الاكتساب Acquisition phase وهي المرحلة التي يكتسب فيها المن ما يكفي من الجسيمات الفايروسية ليكون قادرا على نقلها بعد تغذيته على النبات المصاب بما يسمى "بتغذية الاكتساب" Acquisition feed وتسمى المدة اللازمة لإنجازها "مدة تغذية الاكتساب" Acquisition feeding period وهي المدة التي تحتاجها الحشرة لاكتساب الفايروس من أنسجة النبات المصاب (2) فترة الحضانة Incubation period تسمى فترة الكمون Latent period وهي الفترة الزمنية التي يبقى فيها الفايروس في جسم الحشرة الناقلة لتصبح بعدها قادرة على نقله إلى النباتات السليمة وتتباين هذه الفترة حسب نوع العلاقة القائمة بين الفايروس والمن فهي تستغرق مع الفايروسات المحمولة داخليا بين عدة ساعات إلى عدة أيام أما مع الفايروسات المحمولة خارجيا فهي لا تستغرق إلا عدة ثواني (3) فترة الاحتفاظ بالفايروس Retention period وتسمى أيضا فترة النقل Transmission period وهي الفترة التي يبقى فيها الفايروس فعالا في جسم حشرة المن وتكون فيها ناقلة له إلى النباتات السليمة (4) مرحلة تلقيح النبات السليم Inoculation period وهي المرحلة التي تكون فيها الحشرة الناقلة قادرة على تلقيح النبات بالفايروس الذي تحمله ويشار إلى المدة التي تحتاجها الحشرة لتلقيح النبات السليم بأنها "مدة تغذية التلقيح" Inoculation feeding period أما التغذية فتسمى "تغذية النقل أو التلقيح" Transmission or Inoculation period.
إن التسميات المستعملة لوصف هذه العلاقات وضعت من قبل Watson و Roberts سنة 1939 فهما أول من استعمل مصطلح الفايروس الباقي Persistent Virus والفايروس غير الباقي Non-persistent virus لوصف الفايروسات حسب المدة التي تبقى فيها في المن القادر على نقلها بعد اكتسابه لها.
5- نقل الفايروسات ميكانيكيا بحشرات المن
هي علاقة قليلة الأهمية جدا في نقل الفايروسات وليست تخصصية حيث يتم هذا النقل عن طريق جرح الخلايا النباتية ميكانيكيا بسبب حركة المن على سطح النبات ومع أنواع معينة من الفايروسات الثابتة وهي فايروسات موزائيك التبغ (TMV) والموزائيك الاصفر للشلغم (TYMV) وموزائيك الفاصوليا الجنوبي (SBMV) وهو نقل لم يلاحظ طبيعيا في البيئة ولكنه سجل تجريبيا مع الفايروسات المذكورة الموجودة بتراكيز عالية في النباتات والتي تمتاز بثباتيتها العالية علما بأن هذه الفايروسات لا تنقل بالمن طبيعيا، وتمخضت التجارب التي أجريت في هذا المجال على فايروس موزائيك التبغ عن النتائج التالية (1) لا يمكن الحشرات المن أن تنقل هذا الفايروس برماحها (2) أمكن نقل الفايروس تجريبيا بعمل جروح دقيقة حصلت نتيجة زحف المن على أسطح الأوراق (3) تمكنت حشرات المن من ابتلاع جسيمات الفايروس من النباتات المصابة مباشرة وكذلك من خلال الأغشية النفاذة الصناعية ثم طرحها مع فضلاتها وهي فعالة قادرة على الإصابة (4) لا يثبط لعاب المن الإصابة بالفايروس عند خلطهما معا ثم تلقيح النباتات الكاشفة ميكانيكيا بالخليط (5) أصبحت حشرات المن قادرة على اكتساب الفايروس من خلال الأغشية وذلك عند خلط المحلول الفايروسي النقي مع مركب الأورثينون المتعدد Poly Orthinone وكلوريد البوتاسيوم وفسر ذلك بأن هذا المركب جعل من الخلايا المخترقة بالرمح حساسة للإصابة أو أنه سهل احتفاظ رمح المن بالفايروس.
2. النقل بقفازات الأوراق
قفازات الأوراق Leafhoppers هي حشرات ثاقبة ماصة تنتمي كل أنواعها الناقلة للفايروسات إلى تحت عائلتين هما Agallinae والتي تتغذى أنواعها على النباتات العشبية ثنائية الفلقة و Deltacephalinae التي تتغذى أنواعها على النباتات أحادية الفلقة وهما تابعتين لعائلة قفازات الأوراق Cicadellidae وهي العائلة التي تضم 60 تحت عائلة، وبلغت أعداد الأنواع المسجلة لقفازات الأوراق إلى ما يقرب من 15000 نوع تقع في 2000 جنسا إلا أن الأنواع المسجلة ناقلات لفايروسات النبات هي 49 نوعا تنتمي إلى 21 جنسا، (الجدول 2) وهي حشرات ذات دورة حياة بسيطة حيث يفقس البيض عن حورياتNymphs تتغذى بالامتصاص ثم تمر بعدة انسلاخات قبل وصولها لمرحلة البلوغ وقد يكون لها عدة أجيال في السنة وأحيانا جيلا واحدا، وتشتي الأنواع المختلفة منها إما بشكل بيض أو باللغات أو أشكال غير بالغة، ويبين الشكل (11) مقطعا في الرأس والصدر مع التراكيب الرئيسية فيهما، تتكون الغدة اللعابية وهي العضو المهم لنقل الفايروسات بالقفازات من غدة رئيسية رباعية الفصوص وغدة مساعدة، أما الجهاز الهضمي فيتكون من ثلاث مناطق هي المعي الامامي Foregut والمعي الاوسط Midgut والمعي الخلفي Hindgut، (الشكل 12). إن السلوك التغذوي للقفاز مماثل لسلوك المن حيث يحيط الغشاء اللعابي بأجزاء الفم الذي يخترق أنسجة النبات وصولا إلى اللحاء.

الشكل (11): مقطع في راس حشرة قفاز الاوراق Graminella nigiformis يبين القناة الهضمية الامامية والتراكيب المرافقة مقدمة التجويف الفمي Precebarium الذي يحوي المتحسسات الكيماوية ومحقنة اللعاب Salivary syringe.
مفاتيح الشكل: (1) الصمام المريئي (2) المريء (3) المخيخ Tentorial bar (4) الدماغ (5) العقدة تحت المريئية (6) البلعوم (7) العضلة المتحكمة بالتجويف الفمي Cibarial dilateral (8) muscle التجويف الفموي Cebarium الحاوي على المتحسسات الكيماوية (9) العضلة القابضة Piston retractor muscle (10) الشفة العليا Labrum (11) الرمح (12) الشفة السفلى Labium.
الشكل مقتبس من Hull (2002).
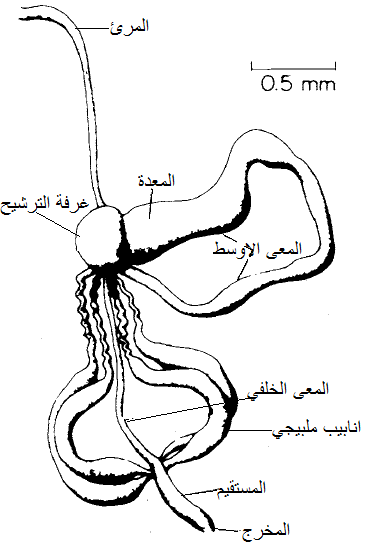
الشكل (12): الجهاز الهضمي للقفاز Agallia comstricta.
الشكل مقتبس من Hull 2002)).
سجلت العلاقات شبه الباقية والباقية بنوعيها المتضاعفة وغير المتضاعفة مع قفازات الأوراق باستثناء العلاقة غير الباقية التي لم تسجل مع أي من القفازات الناقلة للفايروسات وإن العلاقة بين القفازات والناقلات هي متخصصة جدا إذ لا ينقل الفايروس إلا بنوع معين واحد من القفازات أو بعدد قليل من الأنواع في أحسن الأحوال.
أ- العلاقة شبه الباقية
تتماثل مواصفات هذه العلاقة مع مثيلتها في المن ولم تسجل هذه العلاقة إلا مع الفايروسات التابعة لجنس Waikavirus، عائلة Sequiviridae اذ لوحظت جسيمات فايروسي التقزم الاصفر للذرة (MCDV) مرتبطة بالبطانة الكيوتكلية للقناة الهضمية الأمامية لقفاز الأوراق الناقل Graminella nigrifrons وأن الوقت الأدنى لاكتسابه هو بحدود 15 دقيقة ويفقده القفاز خلال أقل من 24 ساعة من اكتسابه عند درجة 25م ولكنه يحتفظ به لعدة أيام عند درجات الحرارة المنخفضة كما تفقده الحوريات بعد الانسلاخ وتنقله الذكور والإناث على حد سواء إلا أن الأخيرة هي الأكثر كفاءة في النقل، ووجدت جسيماته مرتبطة بالبطانة الكيوتكلية للمعي الأمامي والتجويف الفمي الامامي Precibarium والتجويف الفمي الحاوي على المتحسسات الكيماوية Cibarium وكذلك في البلعوم والمريء، حيث يقوم القفاز بقذف الغذاء من المعي الامامي إلى خارج الجسم ويعتقد أن هذا السلوك هو الذي يسبب نقل الفايروس أثناء تغذية القفاز بالية الابتلاع والقذف Ingestion-egestion mechanism، وسجل نقل فايروس التدهور الكروي للرز (RTSV) بقفاز الأوراق Nephotettix virescens مرتبطا بالبطانة الكيوتكلية للقناة الهضمية الأمامية وبوقت أدنى للاكتساب قدره 5-30 دقيقة ويرتبط نقل هذا الفيروس بالقفاز حيويا بنقل فايروس التدهور الباسيلي للرز (RTBV) أيضا عند وجودهما معا في النباتات حيث يسببان مرض تدهور الرز Rice tungro disease، وهناك أدلة على وجود بروتين مساعد مماثل لما سبق ذكره مع فايروسات Potyviruses لنقل الفايروسين المذكورين بالقفاز.
ب- العلاقة الباقية الدوارة
تتماثل مواصفات النقل الباقي بقفازات الأوراق تماما مع مواصفاته في المن، وتعود أنواع الفايروسات المنقولة بالقفازات بهذه الطريقة لجنسي Mastrevirus وCurtovirus ، عائلة Geminiviridae ويصل مستوى التخصص إلى أعلى مراتبه في هذه العلاقة حيث يتخصص نوع من القفاز لكل فايروس ومثال ذلك فإن الناقلات الوحيدة لفايروسات تجعد قمة البنجر (BCTV) وتخطط الذرة (MSV) وتقزم الحنطة (WDV) هي قفازات الاوراق Circulifer tenellus و Cicadulina mbila و Psammotettix alienus على التوالي وتتراوح فترة اكتساب الفايروسات بين عدة دقائق - ساعة وكلما زادت هذه الفترة كلما زادت سرعة النقل ومدة الاحتفاظ بالفايروس في جسم القفاز، أما فترة الحضانة فتصل إلى ما يقرب من 24 ساعة، وتمت متابعة حركة جسيمات فايروس تخطط الذرة (MSV) في ناقله فوجد أنه يتحرك من غرفة الترشيح Filter Chamber إلى الخلايا الامامية للمعي الأوسط Ventriculus وذلك بواسطة "الية الابتلاع الميسرة بالمستقبل" Receptor mediated endocytosis وان المستقبل الذي يعمل في الغدة اللعابية يختلف عن ذلك الموجود في المعي الأوسط وأن الكابسيد هو العامل المحدد للنقل التخصصي بالناقلات.
ج- العلاقة المتضاعفة
تتماثل هذه العلاقة في صفاتها مع مثيلتها في المن حدث تتضاعف الفايروسات المنقولة بهذه الطريقة في القفازات، وسجلت هذه العلاقة في فايروسات تعود لأربعة أجناس هي Phytoreovirus، عائلة Reoviridae و Cytorthabdovirus و Nucleorhabdovirus، عائلة Rhabdoviridae و Marafivirus غير المنتمي لعائلة وتحتاج هذه الفايروسات إلى فترة حضانة طويلة جدا تصل إلى 15 يوم ولا يعرف بالضبط سبب طول هذه الفترة ولكن يعتقد أنها لازمة لتضاعف الفايروس داخل الناقل لكي يصبح مهيئا للنقل، وهناك أدلة على أن هذه الفايروسات تسلك مسارا آخر في حركتها داخل الناقل فضلا عن الجهاز الهضمي والغدد اللعابية وهو المسار عبر الجهاز العصبي للحشرة، (الشكل 13). توجد العديد من الفايروسات المهمة اقتصاديا والتي تنقل بهذه الطريقة ومنها فايروسات عائلة Reoviridae وهي أولى الفايروسات التي ثبت تضاعفها في القفازات بواسطة نوعين من الدراسات التجريبية وهما (1) اختبار النقل بالبيض Transoviral transmission حيث أن فقس البيض عن حوريات حاملة للفايروس وناقلة له هو دليل على تضاعف ذلك الفايروس في القفاز (2) اختبار النقل المتسلسل للفايروس في أفراد متعاقبة من الحشرات حيث تحقن الحشرة الأولى بتركيز معلوم من الجسيمات الفايروسية بواسطة أبره دقيقة في منطقة البطن وتترك لتتغذى لفترة كافية على نبات منيع لذلك الفايروس تكفي لتضاعفه داخلها ثم تسحق الحشرة ويؤخذ مستخلصها الذي يخفف إلى تخفيف معلوم للجسيمات الفايروسية ويحقن في حشرة ثانية وهكذا تستمر العملية مع حشرات أخرى ولغاية الحشرة الأخيرة ولتكن العاشرة مثلا والتي سيبقى تركيز الفايروس فيها عاليا في مستخلصها ومقاربا لتركيزه في الحشرة الأولى التي تم حقنها به إن كان متضاعفا فيها ولو لم يكن كذلك لأنخفض تركيزه في الحشرة الأخيرة إلى 1010 ويتم إثبات ذلك من خلال فحص عينات من دم الحشرات بالمجهر الالكتروني وكذلك قدرة الحشرة الأخيرة على إصابة النبات بالفايروس عند تغذيتها عليه إذ لو كان الفايروس غير متضاعفا فيها لفشلت تلك الحشرة في نقله بسبب تخفيفه العالي جدا في دمها. ولقد تم الكشف عن وجود بروتين أطلق عليه الرمز P2 والذي يربط الجسيمات الفايروسية بخلايا القناة الهضمية للحشرة كما سجل وجود بروتين أخر في الأغشية الخلوية للقفاز Niloparvata lugens يعتقد أنه المستقبل لجسيمات فايروس التبقع الحلقي للبابايا (PRSV) ودليل ذلك هو إمكانية اعاقة نقل هذا الفايروس بواسطة بروتين الأشواك الفايروسية Viral Spike protein الذي بحجم 39 كيلو دالتون الذي تم تخليقه في البكتريا وإطعامه للقفاز الناقل قبل تغذيته على نباتات الرز المصابة بالفايروس حيث أدى إلى تثبيط قدرة القفاز على نقل الفايروس المذكور نتيجة ارتباطه به، وأثيرت تساؤلات حول احتمال التأثير المرضي لهذه الفايروسات على الحشرات الناقلة ولكن لم تثبت الدراسة التي اجريت علي فايروس ريادو فينو الذرة (MRFV) حصول تأثير مرضي يسببه علي قفاز الأوراق Dalbulus maidis رغم أنه ينتشر في أعضاء الناقل.

الشكل (13): نموذج للنقل الدوار لفايروسات الرابدو Rhabdoviruses بواسطة قفاز الأوراق في حالة تغذية على النبات. يكتسب القفاز الفريونات بواسطة الرمح من أوعية لحاء النبات لتنتقل إلى القناة الهضمية الوسطى حيث تغزو الخلايا الطلائية وتنتقل منها إلى الجهاز العصبي للحشرة والقصيبات والهيمولمف ثم تنتشر في كل أنسجة الحشرة لتصل إلى الغدة اللعابية والجهاز التناسلي، تتراكم الفريونات بكمية كبيرة في الغدة اللعابية لتتحرر منه إلى القناة اللعابية وتقذف مع اللعاب في النبات الجديد.
الشكل مقتبس من Jacksonوآخرون (2008).
يتأثر نقل وتضاعف الفايروسات في قفازات الأوراق بعوامل عدة هي (1) عمر الناقل وطوره عند اكتسابه للفايروس فالحوريات هي الأكثر كفاءة فيما تقل كفاءة البالغات في نقل الفايروسات كلما تقدمت في العمر، حيث لوحظ أنه عند تغذية القفاز A.constricta على النباتات المصابة بفايروس التورم الجرحي (WTV) فأن جسيماته وصلت إلى الدم والغدد اللعابية بعد حوالي 30 يوما من الاكتساب وذلك في 50% من الحوريات بينما لم يلاحظ ذلك الا في 5% فقط من البالغات في نفس الأنسجة المذكورة ويفسر ذلك بانه بزيادة عمر الحشرة تصبح الامعاء أكثر مقاومة للإصابة وقد لا يكون الفايروس الموجود في غرفة الترشيح في البالغات قادرا على العبور إلى تجويف الجسم (2) درجات الحرارة والتي تؤثر على انتشار الفايروسات في جسم القفاز فوجد أن تعريض القفاز A.constricta لدرجات حرارية عالية نسبيا تصل إلى 36 م أعاق انتشار فايروس التورم الجرحي من الأمعاء إلى الدم فالغدد اللعابية ثم اختفت الجسيمات بعد ستة أيام من غرفة الترشيح في 60% من الحوريات كما تؤثر الحرارة على النقل بالبيض اذ تنخفض معدلات النقل عند ارتفاع الحرارة إلى ما يقرب من 33م (3) التباين الوراثي في القفازات إذ تتباين سلالات الحشرة في كفاءة نقلها للفايروسات (4) تغير خواص الفايروسات عند تكرار نقلها في النباتات او عند استمرار بقائها في عائلها النباتي المعمر من دون أن تنقل بالقفاز فقد يسبب ذلك فقدان القفازات لقدرتها على نقل الفايروسات حيث لوحظت هذه الظاهرة مع فايروس التورم الجرحي وبعض فايروسات "الرابدو" Rhabdoviruses .
(3) النقل بقفازات النبات
تنتمي قفازات النبات Planthoppers إلى فوق العائلة Fulgoroidae والتي تضم 20 عائلة إلا أن الأنواع الناقلة لفايروسات النبات تقع فقط في عائلة Delphacidae والتي تنتمي إليها حشرات متخصصة بالتغذية على النباتات أحادية الفلقة وخاصة النجيليات حيث تنقل فايروسات خطرة تصيب الرز والحنطة والذرة، (الشكل 2) و (الجدول 1) والعلاقة بين هذه الحشرات والفايروسات المنقولة بها هي من النوع الباقي المتضاعف ولم تسجل أية علاقة أخرى، وقد سجلت هذه العلاقة مع أنواع من الفايروسات تعود إلى ثلاثة أجناس هي Fijivirus و Oryzavirus ، عائلة Reoviridae و Tenuivirus غير المنتمي الى عائلة والذي تنقل أنواعه بقفازات النبات Delphacid حيث تكتسب خلال 15 دقيقة إلى أربع ساعات مع فترة حضانة تمتد من 4-31 يوما لتبقى بعدها الحشرات محتفظة بهذه الفايروسات طيلة حياتها، أما الفترة التي تحتاجها لتلقيح النبات السليم فهي 30 ثانية إلى عدة دقائق وأحيانا قد تستغرق عدة ساعات، والحوريات هي الأكثر كفاءة في النقل من البالغات والإناث أكثر كفاءة من الذكور، وتغزوا هذه الفايروسات معظم أعضاء وأنسجة الناقل إذ وجدت في الدماغ والأجهزة الهضمية والتنفسية والتكاثرية وفي الغدد اللعابية وأنابيب ملبيجي وعضلات الأرجل والأجسام الدهنية مما يسبب إصابات مرضية معتدلة للحشرات مع خفض مستوى الإخصاب Fecundity وموت البيض وتبكير طور الحورية وقد تحصل أحيانا إصابة حادة للحشرات مسببة موتها بسبب حملها للفايروس.
)4) النقل بالذباب الأبيض
الذباب الأبيض Whiteflies هي حشرات صغيرة نشطة تعود لعائلة Alyerodidae، (الشكل 6) و (الجدول 2) ، تتغذى على اللحاء وتضع بيضها على السطح السفلي للأوراق وذات أجزاء فم ثاقبة ماصة إذ تمتلك رمحا شبيها برمح المن مكونا من حزمة الرمح Stylet bUndle المتكونة من الرمحين الفكيين والرمحين المساعدين والتي تحفظ في التجويف Rostrum، وتتماثل هذه الحشرات في سلوكها التغذوي مع حشرات المن والأنواع المسجلة منها كناقلات للفايروسات النباتية هي حشرات Bemisia tabaci و B. Argentifolia و Trialeurodes abutionea و T. vaporariorum وهي تنقل أنواعا فايروسية تعود إلى ثلاثة أجناس هي Begormovirus ، عائلة Geminiviridae و Crinivirus و Closterovirus ، عائلة Closteroviridae و Carlavirus غير المنتمي لعائلة والتي تنتشر أنواعها في المناطق الاستوائية وشبه الاستوائية والمعتدلة وهي مهمة اقتصاديا وأكثر المحاصيل تأثرا بها هي البقوليات والطماطة والقطن. وتعد حشرة ذبابة القطن البيضاء B. tabaci هي الأكثر دراسة لأهميتها في النشر الوبائي للعديد من الفايروسات وهي تمتلك أربعة أطوار من الحوريات التي ترتبط بشدة بالنبات باستثناء فترة قصيرة في بداية طور النمو وتدخل أحيانا في أنسجة النبات وتستقر بين الخلايا متغذية على العصير النباتي في اللحاء، وان الطور اليرقي الأول هو الوحيد المتحرك ولكنه لا يبتعد كثيرا عن النبات اما البالغات فهي مجنحة نشطة تنتج عدة أجيال في السنة. تعد فايروسات Begomociruses إحدى أهم الأجناس الفايروسية المنقولة بالذباب الأبيض بالطريقة الباقية المتماثلة صفاتها مع مثيلتها في حشرات المن وشوهدت جسيمات هذه الفايروسات في الخلايا الطلائية للأمعاء والغدد اللعابية ويعتقد أن بروتين السمبيونين Symboinin يلعب دورا في ربط جسيمات الفايروسات بأغشية الحشرة ولكن وجد أن فشل نقل فايروس موزائيك الابيطلون (AbMV) ليس بسبب عدم ربط الكابسيد مع هذا البروتين وبذلك فان هذه النتيجة تلقي بظلال من الشك على دور هذا البروتين في النقل. سجلت العديد من الظواهر المعقدة المرافقة لنقل هذه الفايروسات بالذباب الأبيض ومنها بقاء فايروس تجعد واصفرار أوراق الطماطة (TYLCV) في ناقله لفترة أطول من المتوقع رغم أنه غير متكاثر فيه فضلا عن وجود أدلة على نقله بالبيض كما أنه ينتقل من الذكور الحاملة له إلى الإناث عند التزاوج ووجد أن كابسيد الفايروس يلعب دورا أساسيا في اكتسابه من قبل الحشرة وتحديدا المنطقة المحصورة بين الحامضين الامينيين رقم 134-129. كما يسبب فايروس تجعد أوراق فول الصويا (SCLV) تأثيرات إمراضية خلوية في بعض أنسجة الناقل ويؤثر أيضا على حيويتها وتكاثرها وهذه الظواهر قد تدل على تكاثره في الحشرة. كما ينقل الذباب الأبيض أيضا العديد من الأنواع التابعة للجنسين Crinivirus و Closterovirus بالطريقة شبه الباقية وبذات الصفات التي سجلت لهذه العلاقة مع المن حيث ترتبط هذه الفايروسات بالمعي الأمامي للحشرات، حيث سجل بقاء فايروس الاصفرار المعدي للخس (LIYV) لمدة ثلاثة أيام في ناقله وهو يشفر بروتينين من بروتينات الكابسيد وهما البروتين الكبير CP و Major protein والبروتين الصغير CPm و Minor protein والأخير هو الضروري لنقل هذا الفايروس بالحشرة، فيما يبقى فيروسي اصفرار الخس (LCV) والاعتلال التقزمي الاصفر للقرعيات (CYSDV) لمدة أربعة وتسعة أيام على التوالي في ناقلاتها.
(5) النقل بحشرات الثربس
تعود حشرات الثربس Thrips إلى رتبة Thysanoptera وهي ذات أجزاء الفم الخادش الماص Rasping sucking mouth parts ومن بين 5000 نوع تقريبا من الثربس فأن هناك عشرة أنواع فقط ناقلة لفايروسات النبات وهي تتبع عائلة Thripidae، (الشكل 2) و (الجدول 1) ومعظم أنواع الثربس هي متعددة التغذية وقادرة على التكاثر في مدى واسع من النباتات، ويعد ثربس البصل Thrips tabaci من الحشرات عالمية الانتشار ويتغذى على ما يقرب من 140 نوعا تتبع 40 عائلة نباتية وهو قادر على التكاثر العذري ويرقاته غير نشطة عادة إلا أن البالغات مجنحة ونشطة جدا ويتغذى بامتصاص محتويات الخلايا تحت البشرة Subepidermal cells، تعيش البالغات ما يقرب من 20 يوما وللثربس عدة أجيال في السنة، أما سلوك الثربس عند التغذية فيتميز بقشط بشرة الورقة بواسطة الفك العلوي والذي يثقب سطح الورقة ثم يتم إدخال اثنين من الفكوك المساعدة في خلية النبات وبواسطتهما يتم امتصاص العصارة النباتية تؤدي أجزاء الفم إلى المعي الأمامي ثم الأوسط المتكون من ثلاث مناطق هي الأمامية والوسطى والخلفية ويبطن الكيوتكل القناتين الأمامية والخلفية ويكون ثخينا غير منفذ أما الوسطى فهي ذات طبقة خلوية طلائية لينة، أما الغدة اللعابية معقدة تتكون من غدتين فصيتين وغدتين أنبوبيتين اللتين ترتبطان بالمنطقة الأمامية للمعي الأوسط، (الشكل 14).
تعود الفايروسات المنقولة بالثربس إلى الجنسTospovirus ، عائلة Bunyaviridae التي تنقل بعض أنواعه بحشرات الثربس بالطريقة الباقية المتكاثرة حيث تكتسب اليرقات هذه الفايروسات فقط فيما تعجز البالغات عن ذلك كما تنخفض كفاءة اليرقات في الاكتساب مع تقدمها في العمر وتصبح البالغات الناتجة من اليرقات حاملة له وقادرة على نقلها طيلة حياتها، وأن أفضل طور لاكتساب هذه الفايروسات هو العمر اليرقي الأول First instar nymph والذي يكتسب الفايروس بفترة تغذية بحدود خمس دقائق إلا أن مدة التلقيح قد تمتد إلى ساعتين ويتأثر ذلك بنوع الناقل والنبات، كما تتباين فترة الحضانة مع تباين الحرارة حيث سجلت فترة حضانة لمدة 84 ساعة عند 27م و 171ساعة عند 20م، ورغم أن تركيز الفايروسات في الإناث هو الأعلى إلا أن الذكور هي الأكثر كفاءة في نقلها وذلك بسبب سلوكها التغذوي. يعد الثربس Frankilinella occidentalis هو الاكثر كفاءة في نقل فايروسات الذبول المبقع للطماطة (TSWV) والتبقع التماوتي للزهرة المجزاعة (INSV) والتبقع الاصفر للطماطة (TCSV) والتبقع الحلقي لفستق الحقل (GRSV)، ووجد أن الأفراد الداكنة اللون Light form للنوع F. Schultzei تنقل فايروسات الذبول المبقع للطماطة والتبقع الاصفر للطماطة والتبقع الحلقي لفستق الحقل بكفاءة عالية فيما تنقلها الأفراد فاتحة اللون Light form بكفاءة منخفضة جدا وأنه إذا ما نقل فايروس الذبول المبقع للطماطة بتكرارية من نبات لأخر دون النقل بالثربس فانه يفقد إمكانية نقله بهذه الحشرة. يبين الشكل (15) العلاقة بين الثربس وفايروسات الجنس Tospovirus حيث يتراكم نيوكليوكابسيد فايروس الذبول المبقع للطماطة مع بروتين غير تركيبي حشري موجود في الغدد اللعابية فضلا عن أنسجة أخرى في يرقات وبالغات الثربس F. occidentalis نتيجة عبوره للعديد من أغشية الناقل بواسطة "الآلية الميسرة بالمستقبل" Receptor mediated mechanism ودليل ذلك أن الحشرة لا تكتسب الفايروس إلا إذا كانت الجسيمة كاملة مغلفة حيث أن هذا الفايروس يمتلك أشواكا خارجية Spikes مكونة من نوعين من الكلايكوبروتينات والتي تبرز من غلاف الفايروس لذا فمن المحتمل أن تكون هذه الاشواك هي موقع ارتباط لمستقبل معين موجود في المعي الأوسط للناقل، ولقد عزل نوعين من البروتينات من الثربس المذكور وهما بحجم 50 و 94 كيلودالتون على التوالي مرتبطان مع الكلايكوبروتين الفايروسي TSWVG2 ولوحظ وجودهما في عدة مواقع من جسم الثربس وذلك عند متابعة حركة الفايروس في اليرقات باستعمال تقانة الصبغ بالفلورة المناعية حيث لوحظت الجسيمات أولا في المنطقة الأمامية للمعي الأوسط وذلك بعد حوالي 24ساعة من الاكتساب ثم زاد وجود الجسيمات في المنطقة ولكنها بقيت مقيدة في الطبقة الطلائية لتلك المنطقة لبعض الوقت ثم انتقلت في المرحلة اليرقية المتأخرة إلى الأنسجة العضلية الطولية والحلقية للقناتين الوسطى والأمامية فيما لوحظ إصابة الغدد اللعابية لأول مرة بعد 72 ساعة من الاكتساب وكذلك إصابة الأربطة الرابطة Ligaments للقناة الوسطى مع الغدد اللعابية ولكن لم يلاحظ وجود جسيمات الفايروس في الدم أو في الصفيحة القاعدية Basal lamina للقناة الوسطى وهذا يشير إلى أن الفايروس لا يصل إلى الغدد اللعابية عن طريق الدم ولكن عبر الأربطة الرابطة للمنطقة الأمامية للمعي الأوسط مع الغدد اللعابية وهذا مسار مختلف عما هو معروف مع الفايروسات الباقية في الحشرات الأخرى، كما وجد أن الفايروس إذا ما اكتسبه الثربس في الطور اليرقي الثاني فإنه نادرا ما يصيب القناة الوسطى وبالتالي لا يصل الفايروس نهائيا إلى الغدد العابية. لم يسجل نقل هذه الفايروسات ببيض الثربس.
تساهم أنواع من حشرات الثربس في النقل غير المباشر لأنواع من الفايروسات التي تعود لأجناس llarvirus و Sobemovirus و Carmovirus وهي فايروسات منقولة بحبوب اللقاح حيث تحمل حشرات الثربس تلك الحبوب وتلقح بها أزهار النباتات السليمة ميكانيكيا خلال تغذيتها على الازهار ولكن لا توجد علاقة حيوية تخصصية بين هذه الفايروسات والثربس.
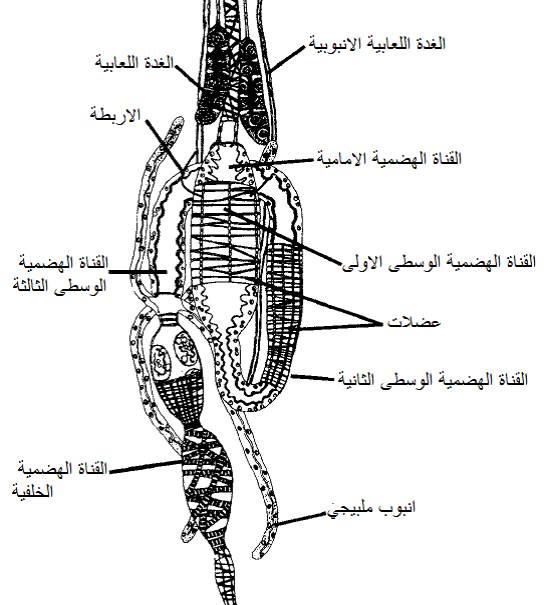
الشكل (14): تركيب القناة الهضمية والاعضاء المرتبطة بها لحشرة الثربس Hercinothrips femoralis.
الشكل مقتبس من Hull (2002).

الشكل (15): مخطط لعلاقة الثربس وفايروسات الجنس Tospovirus حيث يحصل النقل بالبالغات فقط عندما يكتسب الفايروس من قبل اليرقات حيث تنقل يرقات الطور الثاني الفايروسات إلا أن هذه اليرقات هي غير مجنحة ونادرا ما تنتشر بشكل واسع، البالغات هي الطور المنتشر والنشط وهي المسؤولة عن النشر الوبائي لهذه الفايروسات.
الشكل مقتبس من Hull 2002.
(6) النقل بالبق الدقيقي
تنتمي حشرات البق الدقيقي Mealybugs إلى عائلتي Pseudococcidae و Coccoidae، (الشكل 16) و (الجدول 1) وهي ذات أجزاء فم ثاقبة ماصة تتغذى على العصارة النباتية في اللحاء، وهي بطيئة الحركة على النباتات مقارنة بالمن والقفازات لأنها عديمة الأجنحة وتعتمد في حركتها وتنقلها بين النباتات على أرجلها وعلى الرياح فهي تنتقل من نبات لأخر مشيا عند تلامس النباتات حيث تزحف الحوريات وهي الطور الأكثر حركة من البالغات كما يلعب النمل دورا مهما في حملها ونقلها بين النباتات، أما النقل البعيد فيتم بواسطة الرياح، لذلك فهي أقل كفاءة في نقل الفايروسات من الحشرات السابقة المذكورة. وهي حشرات واسعة الانتشار عالميا ويطلق عليها أيضا الحشرات القشرية الكاذبة بسبب عدم امتلاكها للقشور Scales. سجلت العديد من أنواعها ناقلات لعدد من الفايروسات المهمة اقتصاديا وخاصة في المناطق الاستوائية وتشمل الأنواع التابعة لجنس Baonavirus ، عائلة Caulimoviridae وعدد من الأنواع التابعة لجنس Closterovirus، عائلة Closteroviridae ولجنس Vitivirus غير المنتمي لعائلة، وأهم هذه الفايروسات هي الفايروس الثالث المرافق لالتفاف أوراق العنب (3-GLRaV) واصفرار الخس (LCV) والفايروس الأول المرافق لذبول الأناناس المنقول بالبق الدقيقي (1-PMWaV) وفايروس العنب "بي" (GVB) وفايروس العنب "أي" (GVA) وفايروس تضخم أفرع الكاكاو (CSSV) الذي سجلت له ستة أنواع ناقلة وهو يكتسب من قبل الحشرة Pseudococcus nijilansis خلال 20 دقيقة علما بأن الحشرة تحتاج إلى حوالي 16 دقيقة للدخول إلى أنسجة الورقة وتبقى حاملة للفايروس لمدة ثلاث ساعات تقريبا لذا فان العلاقة بينهما تميل نحو العلاقة غير الباقية ويحتمل أن يحمل الفايروس في نهاية الرمح الأمامية، أما الفايروس PMWaV-1 فينقل بحشرة "البق الدقيقي الوردي للأناناس Dysmicoccus brevipes والبق الدقيقي الرمادي للأناناس D. neobrevipes وأن العمر اليرقي الثاني والثالث هما الأكثر كفاءة في اكتسابه من الأعمار الأخرى ومن الإناث، ونظرا لبقاء الفايروس 3-GLRaV في ناقله Planococcus citri لما يقرب من 24 ساعة لذا يحتمل أن تكون العلاقة بينهما شبه باقية.

الشكل (16): حشرة البق الدقيقي
(7) النقل بالبق النباتي
تعود حشرات البق النباتي Bugs إلى عائلتي Piesmatidae و Miridae وتتغذى بالطريقة الثاقبة الماصة بواسطة الرمح، (الشكل 2) ويعتقد أنها ناقلات لأعداد قليلة من الفايروسات ولكن لا زال هناك شك لدى الباحثين بأن هذه الحشرات هي ناقلات ولكن أظهرت بعض البحوث الفردية أن حشرة Cyrtopetis nicotianae ناقلة لفايروسي موزائيك الفاصوليا الجنوبي (SBMV) وتبرقش التبغ الزغبي (VTMOV) ولبعض فايروسات الجنس Sobemovirus وأن الوقت الأدنى للاكتساب هو دقيقة واحدة وهذا دليل على أن العلاقة هي أقرب للعلاقة غير الباقية إلا أن كفاءة النقل ازدادت بزيادة مدة تغذية الاكتساب وهذا دليل على أن العلاقة قد تكون شبه باقية أو ربما باقية، ولا توجد أدلة على تضاعف هذه الفايروسات في هذه الحشرات إلا أنه وجدت جسيمات فايروس تبرقش التبغ الزغبي في أمعاء ودم وبراز الحشرة الناقلة وذلك بعد ستة أيام من الاكتساب ولكنها لم تصل إلى الغدد اللعابية، وتتوفر أدلة على نقل فايروس تجعد أوراق البنجر (BLCV) بالحشرة Piesma quadratum بالطريقة الباقية المتكاثرة ولكن لا توجد أدلة على نقله بالبيض.
(8) النقل بالحشرات القارضة
سجلت أعداد من الحشرات القارضة Biting insects ناقلات للفايروسات في رتبتي مستقيمة الأجنحة Orthoptera وجلدية الأجنحة Dermaptera، (الشكل 2) إلا أن أهم الناقلات تقع في رتبة غمدية الأجنحة (الخنافس) Coleoptera وخاصة في عائلة Chrysomelidae التي تضم الخنافس نباتية التغذية إلا أن 30 نوعا فقط منها سجلت ناقلات لفايروسات النبات، أما في عوائل السوس Curculionidae والدعاسيق Coccinellidae و Meloidae فلم يسجل إلا القليل من الناقلات، (الجدول 1) لا تمتلك الخنافس النباتية غددا لعابية لذلك تتغذى خنافس Chrysomelidae على الأنسجة البارانكيمية الموجودة بين الحزم الوعائية وتترك ثقوبا في الأوراق وقد تتلفها بالكامل عند التغذية الشديدة كما أن لديها سلوك استرجاع الغذاء وقذفه من فمها للخارج Regurgitation ويؤدي ذلك إلى بلل أجزاء فمها بعصير النبات المصاب عند تغذيتها عليه لذلك كان يعتقد سابقا أن الخنافس تنقل الفايروسات نقلا ميكانيكيا بحتا ولكن ثبت خطأ ذلك للأسباب التالية (1) فشل نقل الفايروسات الثابتة ومنها فايروس موزائيك التبغ (TMV) بالخنافس (2) تبقى بعض الفايروسات المنقولة بالخنافس في أجسام ناقلاتها لفترات زمنية طويلة (3) توجد درجة عالية من التخصص بين الفايروسات والخنافس.
تعود الفايروسات المنقولة بالخنافس إلى الأجناس Tymovirus ، عائلة Tymoviridae و Comovirus ، عائلة Comoviridae و Sobemovirus غير المنتمي لعائلة وهي فايروسات ثابتة وتوجد بتراكيز عالية في أنسجة عوائلها النباتية وهي ذات جسيمات أيزومترية بقطر 25-30 نانومتر وتنقل ميكانيكيا وذات مدى عائلي ضيق نسبيا وكذلك تنقل بعدد قليل من أنواع الناقلات. تتباين أنواع الخنافس النباتية في الفترة اللازمة لاكتساب الفايروسات فبعضها يكتسبها بسرعة ومن أول قضمة فيما تحتاج بعض الأنواع إلى فترة تغذية اكتساب تصل إلى 48 ساعة وتزداد كفاءة النقل بزيادة فترة الاكتساب كما تتأثر فترة الاحتفاظ بالفايروس بنوع الفايروس والناقل والظروف البيئية والنبات العائل، إذ وجد أن فترة احتفاظ الخنافس الناقلة لفايروس موزائيك اللوبيا (CPMV) تزداد عند تغذيتها على نباتات اللوبيا المصابة مقارنة بتغذيتها على فول الصويا وذلك بسبب زيادة تركيزه في الأولى، كما وجد أن الخنفساء النباتية Cerotema ruficorniss احتفظت بفايروس الموزائيك المجعد للفاصوليا (BRMV) لتسعة أيام فيما احتفظت به الخنفساء النباتية Diabrolica balteata لثلاثة أيام فقط، كما بقيت الخنافس الخارجة من التشتية حاملة لفايروس تبرقش قرنات البزاليا (BPMV) وقادرة على نقله بعد التشتية وقد يكون هذا دليلا على تضاعفه في الخنافس الناقلة. تظهر بعض الفايروسات بسرعة في دم الحشرات بعد اكتسابها لها كما أصبحت بعض الأفراد حاملة للفايروسات بعد حقنها بها في الدم تجريبيا ولا توجد أدلة على وجود فترة حضانة باستثناء فايروس تبرقش نجيل الإصبع (COMV) المنقول بأنواع من الخنافس Oulema Spp ودليل ذلك هو زيادة معدلات النقل خلال 22 ساعة بعد الاكتساب وفشل الخنافس في النقل بعد الاكتساب مباشرة إلا أنها بدأت بنقله بعد عدة أيام. يعتقد أن نقل الفايروسات بالخنافس هو نقل خارجي أي في أجزاء الفم كما تعد آلية استرجاع الغذاء التي تمارسها الخنافس أثناء التغذية بإعادة قذفها للعصير المسترجع الحامل للفايروس Regurgitant fluid وهي الألية التي تجعل من الخنافس ناقلات للفايروسات. تتحرك كل الفايروسات المنقولة بالخنافس في أوعية الخشب وهي صفة مميزة لهذه الفايروسات تختلف فيها عن بقية الفايروسات التي تعجز في الانتقال عبر هذه الأوعية. لا زالت آلية نقل الفايروسات بالخنافس غير واضحة وبحاجة لمزيد من الدراسات لأنها معقدة وعالية التخصص.
(9) النقل بالحشرات الملقحة للأزهار
توجد العديد من أنواع الحشرات الملقحة للأزهار Pollinating insects والتي يمكن أن تلعب دورا في نقل بعض فايروسات النبات وذلك خلال زيارتها لإزهار النباتات المصابة بالفايروسات لغرض جمع حبوب اللقاح أو امتصاص الرحيق حيث وجد أن فايروس تبرقش أوراق عنب الأحراج (BLMoV) ينقل بحبوب اللقاح التي يحملها النحل وكانت نصف أعداد الشغالات التي أصطيدت تحمل حبوب لقاح مصابة بالفايروس والتي جمعتها من أزهار شجيرات عنب الأحراج.





















