Introduction to Vascular Plants Without Seeds
Vascular plants are traditionally divided into those that do not produce seeds—vascular cryptogams—and those that do- spermatophytes. The former group arose first, and some of its later members were the ancestors of the seed plants.
Beginning about 420 million years ago, certain green algae began to adapt to living on land. Green algae have both marine and fresh water species, and some of the latter must have adapted to the occasional drying of their streams, smaller lakes, and Oceanside mud flats. In environments where long dry periods alternated with long moist periods, the optimal survival strategy probably was formation of dormant, drought-resistant spores. But spore production requires major metabolic conversions; if the dry periods were frequent and temporary, the ability to continue active metabolism by conserving water and adding desiccation would have great selective advantage (Table ). A large, compact, multicellular body would have had a low surface-to-volume ratio and would have automatically retained water better than a small unicellular or filamentous body.

In addition, a waterproofing cuticle would have been selectively advantageous. Evolution of a cuticle may not base been difficult, because fatty acids polymerize automatically in the presence of oxygen, producing a cuticle-like layer. All plants produce fatty acids for their membranes and other metabolic functions, so mutations that permitted some fatty acids to leak to the surface would have resulted in a somewhat waterproof coating and would have had adaptive value. Reproduction would have had to change slightly to coordinate gamete production with periods of moisture, because the sperms had to swim. Also, gamete and spore mother cells would have had to be protected from dryness by one or several layers of cells that formed a jacket around them. Such reproductive modifications resulted in the grouping and protection of spore and gamete mother cells into the sporangia and gametangia characteristic of al embryophytes—relatively massive and protected and having an outer layer of sterile cells
These simple modifications probably allowed the algae not only to survive but also to be metabolically active during short dry periods. But an automatic consequence of much greater advantage was that the algae were safe from predators while they were out of the water Tremendous selective benefit resulted from mutations that enabled the algae to be active for even longer periods out of water. These mutations probably involved increasing the size of the body and the impermeability of the cuticle, but there had to be the simultaneous evolution of stomatal pores and guard cells because a more protective cuticle also prevents the entry of carbon dioxide.
As a truly terrestrial existence became more successful, the mud flats and stream banks would have become crowded, and some plants must have grown over others and shaded them With such shading, the environment became selective for mutations that produced an upright body that could grow into brighter light. This simple change had profound consequences: The strengthening material that evolved, xylem, was also good at conducting water. As phloem evolved, the basal part of the plant that remained in the shade could be nourished. Phloem permitted roots to extend deep into the dark soil, and xylem permitted transport upward of the water and nutrients those roots encountered. The presence of xylem, phloem, and roots freed the plants from their muddy habitats and allowed them to grow anywhere moisture was available near the surface. The presence of the two vascular tissues, necessary for individuals even a few centimeters tall, suddenly gave plants the means to become 100 meters tall, to branch, to form leaves, and to put sporangia high into the air, allowing the spores to be blown great distances, carrying the plant's genes over huge areas, and permitting the colonization of distant habitats.
Vascular tissue, especially phloem, made feasible the evolution of truly heterotrophic tissues—roots, meristems, and organ primordia. Without phloem, each part of a plant can grow and develop only as rapidly as its photosynthesis permits. But the presence of phloem allows mobilization of sugars, minerals, and hormones throughout the entire body and transport to a shoot apical meristem or a group of sporangia, thus permitting a more vigorous, robust growth than could otherwise occur.
One obstacle to the total invasion of land was that plants were still reproductively amphibious. They had terrestrial bodies but aquatic reproduction with swimming sperms. At some point in the life cycle, the environment had to be sufficiently wet that sperms could swim from one gametophyte to another. This requirement is a handicap, but not an insurmountable one: All plants without seeds—nonvascular plants such as mosses and vascular cryptogams such as ferns—still reproduce in this fashion.
The production of pollen and seeds eliminates the need for environmental water £r reproduction. In the line of evolution that led to seed plants, gametophytes became so reduced that they could form completely within the walls of the spores (Fig.). Once this happened, retention of the megaspore and megagametophyte inside the parental sporophyte was feasible. The gametophytes functioned basically as tissue of the sporophyte and benefitted from the land adaptations that had evolved. The microgametophyte, inside the microspore wall, could be transferred by wind to the vicinity of the megasporangium, where it released the sperms into fluids secreted by the megagametophyte. The sperms would swim in this tiny artificial pond and effect fertilization; only the last step of reproduction was still wholly aquatic. In the most advanced seed plants a pollen tube carries nonmotile sperms even further, to the egg itself.
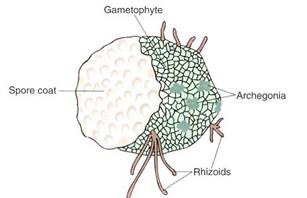
FIGURE :The megagametophyte of Selaginella, which has developed almost completely within the original wall of the megaspore. This is not part of a seed plant but is a necessary first step in seed evolution. Small megagametophytes can be protected and nurtured by their parent sporophyte. By doing so, the gametophyte generation benefits from all those mutations that make the sporophyte adapted for life on land.
Some gametophytes—for example, those of all nonvascular plants—do not need any protection by the parental sporophytes. In most mosses, liverworts, and hornworts, game- tophytes remain rather small and delicate, limiting the entire plant to environments mild enough lor the gametophytes (a plant can live and reproduce only in environments suitable to all essential aspects of its full life cycle), land plants should have been able to develop a life cycle in which gametophytes were just as large, tough, and woody as sporophytes, even having bark, leaves, roots, and so on, but this never happened. the very first land plants did have rather large gametophytes, but in all lines of evolution, gametophytes became simpler and smaller. In the three divisions of nonvascular plants, they are the dominant phase of the life cycle, but nowhere have they achieved the complexity of even the simplest vascular sporophyte. Perhaps in the ngorous terrestrial environment, complex haploid plants and animals cannot compete with diploid plants and animals.
It is common to refer to the features of early species as "primitive" and those of more recently evolved species as "advanced." Unfortunately, in nonscientific English these terms carry judgmental value. "Primitive" usually suggests inferior, inefficient, or poorly functioning, and "advanced" is often inferred to mean superior, efficient, well-adapted, and so on. Used with regard to evolution, these terms may be taken to mean that any plant with advanced features is "better" than one with primitive features. But "better" has no meaning in biology; the proper concept is whether one feature provides more reproductive success, survival value, or selective advantage than another feature. All this depends on the environment and the total biology of the plant. In mild tropical environments, being evergreen is more advantageous than being deciduous because the plants can photosynthesize more, but in environments with stressful winters or summers, being deciduous is more advantageous selectively.
Biologists continually search for neutral words to replace "primitive" and "advanced"; two words that are accurate with regard to evolution and neutral with regard to adaptive value are "relictual" and "derived." Features that were present in the early species of an evolutionary line are described as "original," but the term can really apply only to fossil organisms. Therefore, in a living group that retains many features similar to those of early species, those features are said to be relictual, not original. Features that were not present in the early species but instead evolved later are "derived"; that is, they were derived evolutionarily from the original features. For example, the very first vascular plants evolved from algae, and the bodies of those original species contained only primary growth, only tra- cheids in the xylem, and little sclerenchyma. Reproduction was by spores and free-living gametophytes. Some plants living today have similar features, which are described as relictual. However, many flowering plants have secondary growth, vessels in the xylem, and abundant collenchyma and sclerenchyma, and all produce flowers with dependent gametophytes; these are derived characters. It is tempting to call one set of features primitive and the other advanced, but consider this: Ferns contain all the relictual features just mentioned. Could they be called primitive? They outcompete flowering plants in many environments, perhaps because of their relictual features: By producing no secondary growth and no flowers and by having simple bodies, more of their energy is directed to spore production.





















