























النبات

مواضيع عامة في علم النبات
الجذور - السيقان - الأوراق
النباتات الوعائية واللاوعائية
البذور (مغطاة البذور - عاريات البذور)
الطحالب
النباتات الطبية
الحيوان
مواضيع عامة في علم الحيوان
علم التشريح
التنوع الإحيائي
البايلوجيا الخلوية
الأحياء المجهرية
البكتيريا
الفطريات
الطفيليات
الفايروسات
علم الأمراض
الاورام
الامراض الوراثية
الامراض المناعية
الامراض المدارية
اضطرابات الدورة الدموية
مواضيع عامة في علم الامراض
الحشرات
التقانة الإحيائية
مواضيع عامة في التقانة الإحيائية
التقنية الحيوية المكروبية
التقنية الحيوية والميكروبات
الفعاليات الحيوية
وراثة الاحياء المجهرية
تصنيف الاحياء المجهرية
الاحياء المجهرية في الطبيعة
أيض الاجهاد
التقنية الحيوية والبيئة
التقنية الحيوية والطب
التقنية الحيوية والزراعة
التقنية الحيوية والصناعة
التقنية الحيوية والطاقة
البحار والطحالب الصغيرة
عزل البروتين
هندسة الجينات
التقنية الحياتية النانوية
مفاهيم التقنية الحيوية النانوية
التراكيب النانوية والمجاهر المستخدمة في رؤيتها
تصنيع وتخليق المواد النانوية
تطبيقات التقنية النانوية والحيوية النانوية
الرقائق والمتحسسات الحيوية
المصفوفات المجهرية وحاسوب الدنا
اللقاحات
البيئة والتلوث
علم الأجنة
اعضاء التكاثر وتشكل الاعراس
الاخصاب
التشطر
العصيبة وتشكل الجسيدات
تشكل اللواحق الجنينية
تكون المعيدة وظهور الطبقات الجنينية
مقدمة لعلم الاجنة
الأحياء الجزيئي
مواضيع عامة في الاحياء الجزيئي
علم وظائف الأعضاء
الغدد
مواضيع عامة في الغدد
الغدد الصم و هرموناتها
الجسم تحت السريري
الغدة النخامية
الغدة الكظرية
الغدة التناسلية
الغدة الدرقية والجار الدرقية
الغدة البنكرياسية
الغدة الصنوبرية
مواضيع عامة في علم وظائف الاعضاء
الخلية الحيوانية
الجهاز العصبي
أعضاء الحس
الجهاز العضلي
السوائل الجسمية
الجهاز الدوري والليمف
الجهاز التنفسي
الجهاز الهضمي
الجهاز البولي
المضادات الميكروبية
مواضيع عامة في المضادات الميكروبية
مضادات البكتيريا
مضادات الفطريات
مضادات الطفيليات
مضادات الفايروسات
علم الخلية
الوراثة
الأحياء العامة
المناعة
التحليلات المرضية
الكيمياء الحيوية
مواضيع متنوعة أخرى
الانزيمات

Origin and Evolution of Nonvascular Plants
 المؤلف:
AN INTRODUCTION TO PLANT BIOLOGY-1998
المؤلف:
AN INTRODUCTION TO PLANT BIOLOGY-1998
 المصدر:
JAMES D. MAUSETH
المصدر:
JAMES D. MAUSETH
 الجزء والصفحة:
الجزء والصفحة:
 15-11-2016
15-11-2016
 2665
2665




+
-
20
Origin and Evolution of Nonvascular Plants
No consensus exists about the origin of nonvascular plants; the sparse fossil evidence can be interpreted in various ways. A variety of hypotheses have been offered, but many botanists believe that insufficient information is available to form any firm opinion. Two frequently considered hypotheses are (1) that nonvascular plants evolved from early vascular plants, the rhyniophytes, by becoming simpler and reduced and by losing fair vascular tissues, and (2) that nonvascular plants are not related to vascular plants at all, but rather evolved from green algae separately and at a different time than the rhyniophytes. A third emerging hypothesis suggests that at least some nonvascular plants evolved from vascular ancestors by simplification, but the divergence between the two groups started even earlier than the rhyniophytes; that is, some nonvascular plants evolved from the same group that later gave rise to the rhyniophytes.
The last hypothesis centers on living hornworts like Anthoceros and fossil plants called Horneophyton. The sporophytes of Horneophyton were naked axes that branched dichotomously and were up to 20 cm tall (Fig. ). They had stomata and terminal sporangia. Each sporangium had a short columella, and just as in hornworts, the sporogenous tissue surrounded the columella on all sides and the top. The stem base was swollen, similar to a hornwort foot, except that it bore rhizoids.
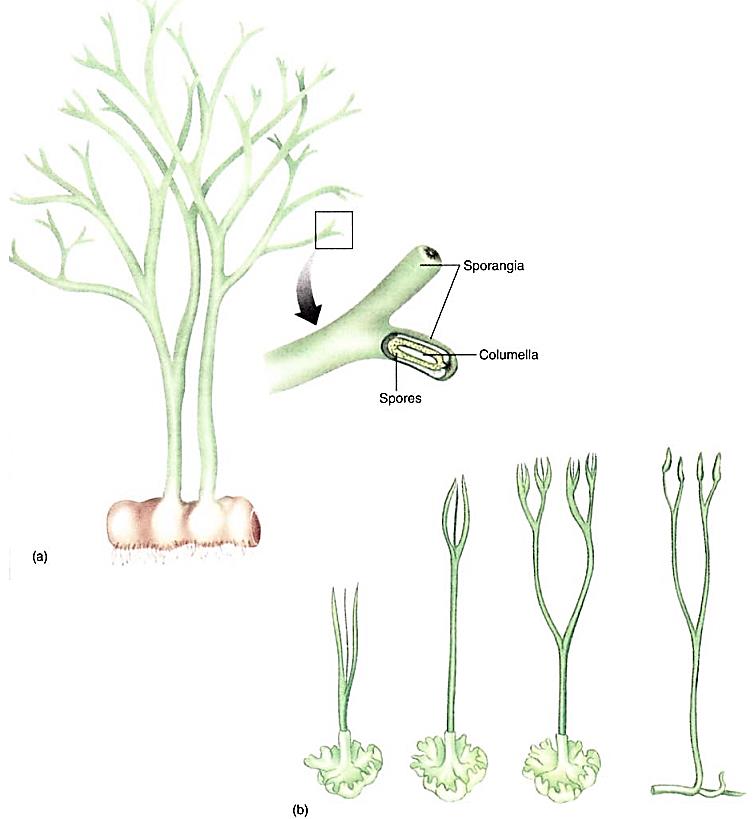
FIGURE : (a) Reconstruction of the extinct plant Horneophyton and its terminal sporangia. (b) At first, it was postulated that algae gave rise to land plants whose life cycle was dominated by the gametophyte and whose sporophyte was small and dependent, hemiparasitic on the gametophyte (as drawn on the left). It was postulated that this evolved into the vascular plants as the sporophyte became larger and more elaborate while the gametophyte became reduced (as drawn on the right). This hypothesis has been rejected and it is now suspected that perhaps the hornworts evolved by the reverse process, reading this series from right to left.
The similarities between Anthoceros and Horneophyton were recognized early in this century, and it was suggested then that vascular plants may have arisen by a process in which (1) algae gave rise to hornworts similar to Anthoceros, with small sporophytes that depended on thalloid gametophytes; (2) these species in turn evolved into Horneophyton- like plants as the gametophytes became simpler and shorter-lived while the sporophytes became taller and more active photosynthetically (Fig. 22.33b, from left to right); (3) finally, the gametophyte was very small and temporary, and the sporophyte was an independent, free-living plant.
This idea is not accepted by many botanists today for several reasons, but it is possible to think about these transitions in reverse—that hornworts evolved from species like
Horneophyton by means of an elaboration of the gametophyte and a reduction of the sporophyte. In the earliest true land plants, gametophytes were not thalloid like those of Anthoceros, but rather had upright stems; there appears to have been an alternation of isomorphic generations. Early true plants appear to have been a group of species that resembled each other but had already diverged to the point that some had sporangia without a columella and others had sporangia with a short, incomplete hornwort-type of columella .
The first group apparently gave rise to the vascular plants, with sporophytes becoming larger and gametophytes becoming smaller. The second group might have been the ancestors to the hornworts, with the gametophytes becoming the dominant phase of the life cycle. If so, vascular plants and hornworts are somewhat related, but the divergence between the two groups would have started very early, even before the full land plant syndrome had evolved. The presence of just one chloroplast per cell and the unusual formation of gametangia can be interpreted as supporting the theory of a completely separate origin in the green algae. Any common, non-algal ancestor of hornworts and vascular plants must have existed very early, before many features were established definitively.
Only the origin of Anthoceros has been discussed, not that of nonvascular plants in general. The few available fossils of nonvascular plants do not support the idea that mosses and liverworts are closely related to hornworts. Mosslike plants do not appear in the M record until the Carboniferous Period, about 300 million years ago, about 100 million years after the hypothesized origin of hornworts. By the Permian Period, fossil mosses was as complex morphologically as living ones today; fossils with the cell arrangement pattern of Sphagnum, called Protosphagnum, occurred about 260 million years ago. Most fossils found in rocks 60 million years old are assigned to modern, extant genera.
The earliest liverwort fossils are much older than those of mosses; Pallavicinites devonicus, as its species name indicates, lived in the Devonian Period (400 million years ago), not long after Horneophyton. Full liverwort features existed by this early time. A few million years later, in the Carboniferous Period, leafy liverworts in the form of Hepaticites kidstoni had evolved; this species is very much like a living liverwort, Treubia. Indeed, the earliest fossils of both groups are well developed as either mosses or liverworts; no known fossil is being interpreted as an ancestor to both. Consequently, it seems prudent to favor the hypothesis that the nonvascular plants are polyphyletic, having had separate origins and having been distinct, well-defined groups before the end of the Paleozoic Era.
 الاكثر قراءة في النباتات الوعائية واللاوعائية
الاكثر قراءة في النباتات الوعائية واللاوعائية
 اخر الاخبار
اخر الاخبار
اخبار العتبة العباسية المقدسة

الآخبار الصحية


مواضيع ذات صلة
 قسم الشؤون الفكرية يصدر كتاباً يوثق تاريخ السدانة في العتبة العباسية المقدسة
قسم الشؤون الفكرية يصدر كتاباً يوثق تاريخ السدانة في العتبة العباسية المقدسة "المهمة".. إصدار قصصي يوثّق القصص الفائزة في مسابقة فتوى الدفاع المقدسة للقصة القصيرة
"المهمة".. إصدار قصصي يوثّق القصص الفائزة في مسابقة فتوى الدفاع المقدسة للقصة القصيرة (نوافذ).. إصدار أدبي يوثق القصص الفائزة في مسابقة الإمام العسكري (عليه السلام)
(نوافذ).. إصدار أدبي يوثق القصص الفائزة في مسابقة الإمام العسكري (عليه السلام)



















