























النبات

مواضيع عامة في علم النبات
الجذور - السيقان - الأوراق
النباتات الوعائية واللاوعائية
البذور (مغطاة البذور - عاريات البذور)
الطحالب
النباتات الطبية
الحيوان
مواضيع عامة في علم الحيوان
علم التشريح
التنوع الإحيائي
البايلوجيا الخلوية
الأحياء المجهرية
البكتيريا
الفطريات
الطفيليات
الفايروسات
علم الأمراض
الاورام
الامراض الوراثية
الامراض المناعية
الامراض المدارية
اضطرابات الدورة الدموية
مواضيع عامة في علم الامراض
الحشرات
التقانة الإحيائية
مواضيع عامة في التقانة الإحيائية
التقنية الحيوية المكروبية
التقنية الحيوية والميكروبات
الفعاليات الحيوية
وراثة الاحياء المجهرية
تصنيف الاحياء المجهرية
الاحياء المجهرية في الطبيعة
أيض الاجهاد
التقنية الحيوية والبيئة
التقنية الحيوية والطب
التقنية الحيوية والزراعة
التقنية الحيوية والصناعة
التقنية الحيوية والطاقة
البحار والطحالب الصغيرة
عزل البروتين
هندسة الجينات
التقنية الحياتية النانوية
مفاهيم التقنية الحيوية النانوية
التراكيب النانوية والمجاهر المستخدمة في رؤيتها
تصنيع وتخليق المواد النانوية
تطبيقات التقنية النانوية والحيوية النانوية
الرقائق والمتحسسات الحيوية
المصفوفات المجهرية وحاسوب الدنا
اللقاحات
البيئة والتلوث
علم الأجنة
اعضاء التكاثر وتشكل الاعراس
الاخصاب
التشطر
العصيبة وتشكل الجسيدات
تشكل اللواحق الجنينية
تكون المعيدة وظهور الطبقات الجنينية
مقدمة لعلم الاجنة
الأحياء الجزيئي
مواضيع عامة في الاحياء الجزيئي
علم وظائف الأعضاء
الغدد
مواضيع عامة في الغدد
الغدد الصم و هرموناتها
الجسم تحت السريري
الغدة النخامية
الغدة الكظرية
الغدة التناسلية
الغدة الدرقية والجار الدرقية
الغدة البنكرياسية
الغدة الصنوبرية
مواضيع عامة في علم وظائف الاعضاء
الخلية الحيوانية
الجهاز العصبي
أعضاء الحس
الجهاز العضلي
السوائل الجسمية
الجهاز الدوري والليمف
الجهاز التنفسي
الجهاز الهضمي
الجهاز البولي
المضادات الميكروبية
مواضيع عامة في المضادات الميكروبية
مضادات البكتيريا
مضادات الفطريات
مضادات الطفيليات
مضادات الفايروسات
علم الخلية
الوراثة
الأحياء العامة
المناعة
التحليلات المرضية
الكيمياء الحيوية
مواضيع متنوعة أخرى
الانزيمات

Internal Structure of Foliage Leaves
 المؤلف:
AN INTRODUCTION TO PLANT BIOLOGY-1998
المؤلف:
AN INTRODUCTION TO PLANT BIOLOGY-1998
 المصدر:
JAMES D. MAUSETH
المصدر:
JAMES D. MAUSETH
 الجزء والصفحة:
الجزء والصفحة:
 13-11-2016
13-11-2016
 7248
7248




+
-
20
Internal Structure of Foliage Leaves
EPIDERMIS
Flat, thin foliage leaves (optimal for light interception) have a large surface area through which water can be lost. Water loss through the epidermis is called transpiration, and it is a serious problem if the soil is so dry that roots cannot replace the water lost from the leaves. The epidermis must be reasonably waterproof but simultaneously translucent, and it must allow entry of carbon dioxide. The structure of the leaf and stem epidermis is basically quite similar, consisting of a large percentage of flat, tabular (shaped like paving stones), ordinary epidermal cells; guard cells and trichomes (either glandular or nonglan- dular) may be abundant. However, the dorsiventral nature of the leaf causes the upper and lower epidermis to exist in significantly different microclimates (Fig. 1).
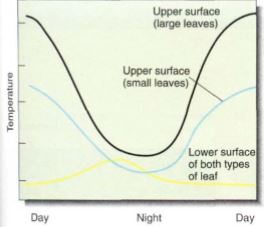
FIGURE 1: At night, both the upper and lower surfaces of a leaf are close to air temperature; some loss of water causes evaporative cooling. After the sun rises, the upper surface rapidly becomes warmer than the surrounding air; in large simple leaves this is much more striking than in small compound leaves.
On a clear, sunny day, a leaf is usually warmer than the surrounding air, so it heats the air and convection currents rise from it. If stomata in the upper epidermis are open and losing water, the water molecules are swept away by this convection. However, air tends to be trapped on the underside of a leaf, so water loss from the stomata there is not so great. The molecules are trapped in quiet air and may diffuse back into the stomata. In most leaves, the number of stomata per square centimeter is much greater in the lower than in the upper epidermis; in many species, no stomata at all are found in the upper epidermis (Table).
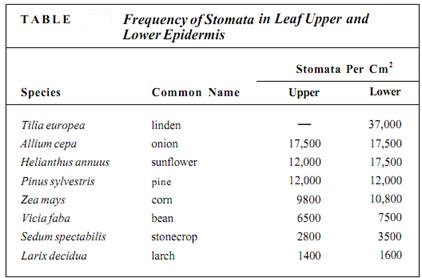
Such a unilateral distribution of stomata has other beneficial consequences. Air-borne spores of fungi and bacteria are continually landing on leaves. Rye leaves, for example, typically have 10,000 fungal spores per square centimeter, but most are on the exposed upper surface, where stomata are few, so few fungi are able to penetrate the leaf.
Leaf stomata are frequently sunken into epidermal cavities that create a small region of nonmoving air. In oleander, the lower leaf epidermis contains numerous crypts (areas where the epidermis is depressed into the leaf), and stomata and trichomes are abundant in the epidermis that lines the crypts (Fig. 2). Both the recessed nature of the crypt and the presence of trichomes keep air in the crypt extremely quiet; many water molecules that diffuse out of open stomata pores bounce around the crypt and re-enter the stomata rather than being blown away.

FIGURE 2 :(a) Oleander (Nerium oleander) leaves have stomatal crypts filled with trichomes and stomata. This organization minimizes air movement near the stomata, and water molecules that diffuse out of the stoma may re-enter the stoma by random motion. Crypts occur only in the lower surface (X 80). (b) Each of the stomata of this grass (Elytrostrachys) is overarched by four protrusions from surrounding cells. This too reduces air movement near the stoma (X 940). (c) In this bamboo (Chusqua), stomata are protected by protrusions and wax rods (X 940). (b and c, Courtesy of L G. Clark and X. Londono, Iowa State University)
Leaf epidermises are often remarkably hairy, and the trichomes affect the leaf biology in numerous ways. They provide some shade on the upper surface of the leaf, deflecting excessive sunlight, an adaptation common in desert plants (Fig. 3). On the lower surface, they prevent rapid air movement and slow water loss from the stomata. In any position, trichomes make walking or chewing difficult for insects, and many glandular trichomes secrete powerful stinging compounds that prevent even large animals from eating the leaf. Insects either do not bother with the leaf or must walk so slowly on it that fey become more vulnerable to their own predators. Conversely, hairs provide excellent footholds against leaf flutter for insects of the appropriate size. Poisonous glandular trichmes in stinging nettle and other species prevent mammals from eating leaf tissue but also protect leaf-borne insects small enough to walk, feed, and reproduce safely between the stinging trichomes.

FIGUREb3: (a) The leaves of lamb's ears (Stachys) are so densely covered with hairs that they often appear white. The hairs greatly reduce the amount of heat that these leaves absorb from sunlight. (b) These plants of Sempervivum are native to high alpine regions where ultraviolet radiation is intense. The hairs of the leaf epidermis reflect part of the ultraviolet radiation away from the plant.
Like stem epidermis, leaf epidermal cells contain a coating of cutin and usually also wax on their outer walls. These retain water and make digestion by fungi difficult. Furthermore, their smooth, slippery surface prevents spores from sticking or allows them to be wasted of by rain.
MESOPHYLL
The ground tissues interior to the leaf epidermis are collectively called the mesophyll (Figs. 4 and 6). Along the upper surface of most leaves is a layer of cells, the palisade parenchyma, which is the main photosynthetic tissue of most plants. Palisade cells are separated slightly so that each cell has most of its surface exposed to the carbon dioxide of the intercellular space. Because carbon dioxide dissolves into cytoplasm slowly, the large surface gives maximum area for dissolution; tightly packed cells could not absorb enough carbon dioxide for efficient photosynthesis (Fig. 5). The palisade parenchyma is oifcn only one layer thick, but in regions with intense, penetrating sunlight, it may be three or four layers thick (Fig. 6).
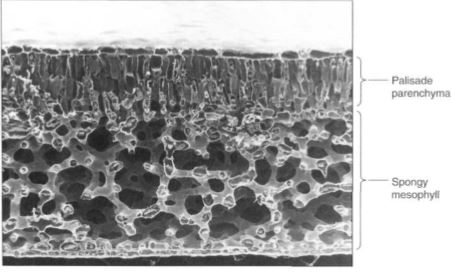
FIGURE 4: This leaf of Laurelia has a layer of palisade parenchyma that is two cells deep, and the spongy mesophyll is an extensive aerenchyma (X 180).
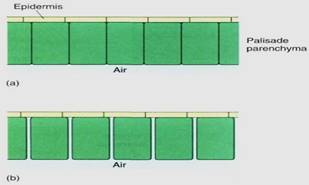
FIGURE 5: (a) If palisade cells are closely packed with no space between them, only the bottoms of the cells are exposed to the carbon dioxide in the spongy mesophyll. (b) With only slight separation of the cells, the volume of the palisade is almost unchanged, but the surface area available is increased enormously; carbon dioxide can be absorbed much more rapidly. Assume that the palisade parenchyma cells are rectangular columns 20 pm X 20 pm X 100 pm long; compute the surface area exposed to CO2 by 100 cells arranged as in (a) versus (b).
In the lower portion of the leaf is the spongy mesophyll—open, loose aerenchyma tissue that permits carbon dioxide to diffuse rapidly away from the stomata into all parts of the leaf's interior (see Figs. 4 and 6). If stomata were surrounded by closely packed cells, a molecule of carbon dioxide might simply bounce off a cell and escape back out of the stomatal pore.
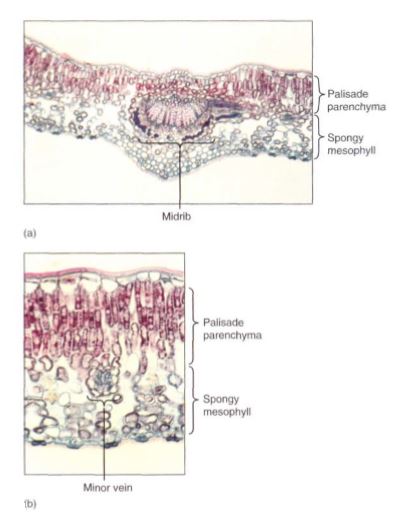
FIGURE 6: (a and b) Leaves of holly (Ilex) have an extremely thick palisade parenchyma consisting of at least three layers of columnar cells. Cells of the lowest palisade layer are relatively widely separated, permitting rapid diffusion of CO2 from the spongy mesophyll to the upper palisade (a, X 50; b, X 150).
Although this arrangement is the most common, some plants have a layer of palisade parenchyma along both surfaces of the leaf; the spongy mesophyll either occurs in the center or is lacking (Fig. 7). The relationship between leaf position and the sun is important: For leaves that are held horizontally (most leaves), the sun is usually overhead, so having palisade parenchyma on the upper surface permits maximum absorption of light and photosynthesis. For plants that hold their leaves vertically (Iris, Gladiolus, Eucalyptus), tab sides are equally illuminated and palisade parenchyma is equally functional on either side.
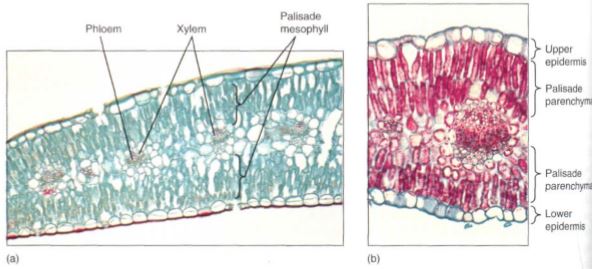
FIGURE 7:Leaves of both carnations (a) and creosote bush (b) have palisade parenchyma along both surfaces. The little spongy mesophyll present is located in the middle of the leaf (a, X 75; b, X 150).
VASCULAR TISSUES
Between the palisade parenchyma and the spongy mesophyll are the vascular tissues. A dicot leaf usually has a large midrib, also called a midvein, from which lateral veins emerge that branch into narrow minor veins .Minor veins are most important for releasing water from xylem and loading sugar into phloem, whereas the midrib and lateral veins are involved mostly in conduction. Vein structure changes with size: The midrib and lateral veins always contain both primary xylem on the upper side and primary phloem on the lower side (Fig. 8). Because they both conduct and support the leaf blade, they may have many fibers arranged as a sheath, called a bundle sheath, around the vascular tissues (Fig.9).
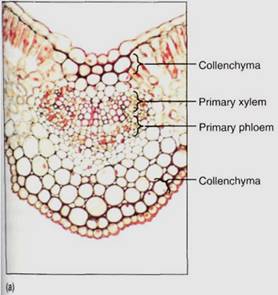
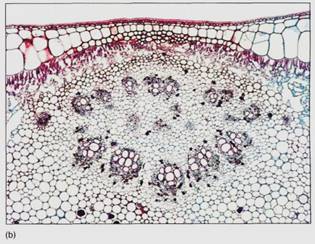
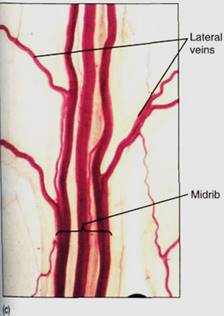
FIGURE 8: Transverse sections through leaf midribs. (a) This midrib of a Ligustrum leaf is rather simple, containing a single vascular bundle. Xylem is present along the top, phloem along the bottom. A small amount of collenchyma occurs on the top and bottom of the midrib (X 150). (b) This midrib of rubber tree has numerous separate bundles embedded in a large mass of mesophyll (X 50). (c) This is a midrib prepared as a leaf clearing ; the three distinct bundles have no interconnections in this region. Each transports to and from a particular portion of the blade (X 15).
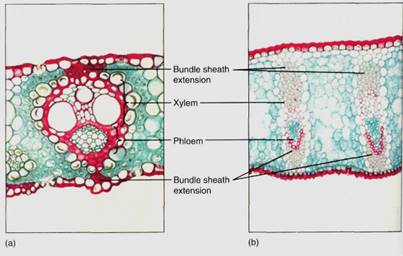
FIGURE 9: :(a) This leaf bundle of sugar cane has bundle sheath extensions contacting both the upper and lower epidermis. Water diffuses from the xylem to the epidermis by capillary action, moving around the surfaces of the fibers (X 100). (b) The leaves of flax have large bundle sheath extensions consisting of many layers of fibers (X 45).
The sheath also makes it difficult for insects to chew into the vascular tissues. Many other types of non-conducting cells such as mucilage, tannin, or starch storage cells may be present in the larger veins. Minor veins are the sites of material exchange with the rest of the mesophyll and must have a large surface area in contact with the palisade and spongy mesophyll; they do not contain fibers or other non-conducting cells whose presence would interrupt this contact (Fig. 10). The endings of the minor veins in some species consist of only xylem and in others only phloem.
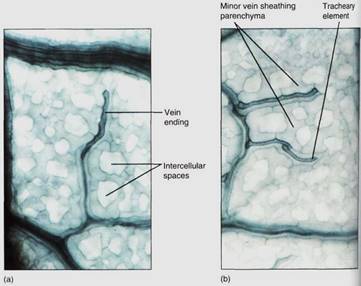
FIGURE 10 (a and b) These leaf veins are minor veins, treated to reveal individual cells. The long dark cells are tracheary elements, and the phloem is out of focus, in line with the xylem. Notice how narrow these veins are: The xylem is only one element wide, so there is maximum surface area for releasing water to the leaf mesophyll. In this species, each minor vein is lined with parenchyma cells that extend outward into the spongy mesophyll (X 250). (Courtesy of N. R Lasten and J. D. Curtis, Iowa State University)
Veins, especially the larger ones, often have a mass of fibers above, below, or both— the bundle sheath extension (see Fig. 9). The fibers help give rigidity to the leaf and are believed to provide an additional means by which water moves from the bundle out to the mesophyll. Apparently water moves by capillary action around the fibers rather than through them.
THE PETIOLE
Although the petiole is technically part of the leaf, it serves as the transition between the stem and the blade. Consequently, the arrangement of tissues differs at the two ends. The epidermis may be similar to that on the lamina but often contains fewer stomata and trichomes. The mesophyll is rather like cortex—somewhat compact and not especially aerenchymatous and considerable collenchyma may be present, providing support for the lamina. Vascular tissues are the most variable; one, three, five, or more vascular bundles, called leaf traces, exit the stem vascular cylinder and diverge toward the petiole (Fig. 11a). They may remain distinct or fuse into a single trace at, near, or in the petiole. In some species, they divide into numerous bundles and 10 or 20 (in large palm leaves, several hundred) may be found in the petiole (Fig. 6.26b). Vascular bundles may either fuse with each other within the petiole or branch further; they can be arranged in a ring, a plate, or a number of other patterns. If the lamina has a strong midrib, most of the petiole bundles fuse together and form the midvein, but other bundles may enter the lamina as small lateral veins.
We do not understand the significance of all the bundle patterns in petioles. Certain patterns may ensure the proper distribution of sugars out of different parts of the lamina, especially of large leaves, into the various bundles of the stem. However, it may be that almost any pattern functions well, so mutations that cause new patterns are not selectively disadvantageous. On the other hand, future studies may show correlations between lamina, petiole, and stem.
In many species, the petiole bears two small flaps of tissue at its base called stipules, which serve various functions. They may protect the shoot apical meristem while the leaf is young and small. In other plants, they are large enough to contribute a significant amount of photosynthesis, but usually when the leaf is mature the stipules are still small and they die early.
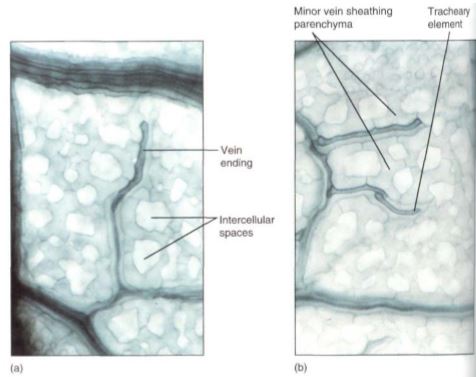
FIGURE 11: (a and b) These leaf veins are minor veins, treated to reveal individual cells. The long dark cells are tracheary elements, and the phloem is out of focus, in line with the xylem. Notice how narrow these veins are: The xylem is only one element wide, so there is maximum surface area for releasing water to the leaf mesophyll. In this species, each minor vein is lined with parenchyma cells that extend outward into the spongy mesophyll (X 250).
 الاكثر قراءة في الجذور - السيقان - الأوراق
الاكثر قراءة في الجذور - السيقان - الأوراق
 اخر الاخبار
اخر الاخبار
اخبار العتبة العباسية المقدسة

الآخبار الصحية


مواضيع ذات صلة
 قسم الشؤون الفكرية يصدر كتاباً يوثق تاريخ السدانة في العتبة العباسية المقدسة
قسم الشؤون الفكرية يصدر كتاباً يوثق تاريخ السدانة في العتبة العباسية المقدسة "المهمة".. إصدار قصصي يوثّق القصص الفائزة في مسابقة فتوى الدفاع المقدسة للقصة القصيرة
"المهمة".. إصدار قصصي يوثّق القصص الفائزة في مسابقة فتوى الدفاع المقدسة للقصة القصيرة (نوافذ).. إصدار أدبي يوثق القصص الفائزة في مسابقة الإمام العسكري (عليه السلام)
(نوافذ).. إصدار أدبي يوثق القصص الفائزة في مسابقة الإمام العسكري (عليه السلام)



















