آخر المواضيع المضافة

 النبات
الحيوان
الأحياء المجهرية
علم الأمراض
التقانة الإحيائية
التقنية الحيوية المكروبية
التقنية الحياتية النانوية
علم الأجنة
الأحياء الجزيئي
علم وظائف الأعضاء
الغدد
المضادات الحيوية
النبات
الحيوان
الأحياء المجهرية
علم الأمراض
التقانة الإحيائية
التقنية الحيوية المكروبية
التقنية الحياتية النانوية
علم الأجنة
الأحياء الجزيئي
علم وظائف الأعضاء
الغدد
المضادات الحيوية| External Structure of Foliage Leaves |


|
|
Read More
Date: 29-10-2015
Date: 21-11-2016
Date: 14-11-2016
|
External Structure of Foliage Leaves
The most obvious function of foliage leaves is photosynthesis, but other functions often taken for granted are just as important. Leaves must not lose excessive amounts of water; they must not allow entry of fungi, bacteria, or epifoliar algae; they must not be so nutritious and delicious to animals that they are a liability to the plant; they must not be such effective sails that the plant is blown over in a mild wind; and they must be cheap enough that the plant spends less carbohydrate building them than it recovers by their photosynthesis. If a foliage leaf fails in any of these aspects, the plant dies and the leafs photosynthesis has been useless. Many of the structural and physiological aspects of leaves that make them waterproof, pathogen resistant, and so on in some way interfere with photosynthesis.
During photosynthesis, leaves absorb carbon dioxide and convert it to carbohydrate by using light energy. Because sunlight comes from one direction at a time, it is best for a portion of the leaf to be flat and wide, allowing maximum exposure to light and maximum surface area for carbon dioxide absorption (Fig. 1). Because chlorophyll absorbs sunlight efficiently, light penetrates only a short distance, so leaves can be quite thin; in a thick layer of chlorenchyma, the lowest cells would be in almost complete darkness, unable to photo- synthesize. This flat, light-harvesting portion is the leaf blade (also called the lamina).
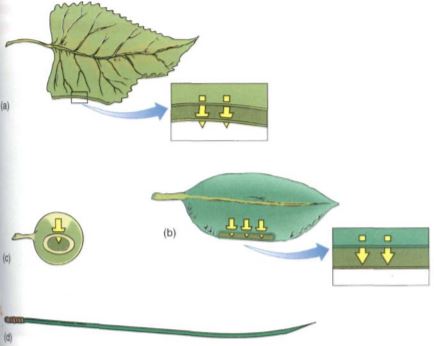
FIGURE 1: Several possible leaf shapes, all with equal volumes. (a) The common type with a thin, flat blade. All cells are exposed to light, there is a large surface area for absorption of CO2, and none of the veins is long. (b) In a thick leaf, light cannot penetrate to the bottom of the leaf and those cells cannot photosynthesize. Relative to the volume, little surface area is available for CO2 absorption, but this helps conserve water. (c) A spherical leaf has maximum internal self-shading, and only a small fraction of the cells receive enough light for photosynthesis. Surface area for gas exchange is minimal. (d) A cylindrical leaf could be thin with a large surface area and no self-shading, as in (a); however, to have the same volume as (a), it would be extremely long, and conduction through the veins might be difficult.
Most leaves have a petiole (stalk) that holds the blade out into the light (Fig. 2), which prevents self-shading of leaf blades by those above them; self-shading would defeat the basic usefulness of the leaf.
The presence of a petiole has other consequences: Long, thin, flexible petioles allow the blade to flutter in wind, cooling the leaf and bringing fresh air to the leaf surface. If the leaf and air are still, carbon dioxide is absorbed from the vicinity of the leaf faster than diffusion can bring more to it, depleting the carbon dioxide and decreasing photosynthesis (Table 1). Leaf flutter also makes it difficult for insects to land on a leaf and knocks off some that have already alighted.

FIGURE 2: The petioles of each leaf of this Astragalus are long enough that the leaves do not shade each other and all are fully exposed to light. The leaves are palmately compound.

If leaves are small or very long and narrow, self-shading is not a problem, and there may be no petiole; the leaf is then sessile instead of petiolate. For example, Aeonium grows in arid, sunny regions, and its fleshy leaves are packed close together (Fig. 3). Because the sunlight is so intense, even with self-shading the leaves receive enough light for adequate photosynthesis. The close packing of these leaves helps trap water molecules and prevent their escape from the plant.

FIGURE 3: Aeonium tabuliforme and its relatives grow in regions of intense sunlight; self-shading is not a problem, but water conservation is. Close packing minimizes water loss from stomata.
In many monocots such as grasses, irises, lilies, and yuccas, foliage leaves have a shape quite different from those of dicots. They tend to be very long and tapering, and self-shading occurs only at the base; most of the blade is well-exposed to light (Figs. 4 and 13). They also typically lack a petiole; instead, the base of the leaf wraps around the stem to fern a leaf sheath. The blade can flex and flutter even without a petiole. Not all monocots have linear, grasslike leaves. Palms, aroids (such as Monstera and Philodendron), and bird- of-paradise plants have what appear to be petioles and laminas, but the leaves of these species are believed to have evolved from grasslike leaves, and the "petiole" is actually a modified portion of the lamina (Fig. 5).

FIGURE 4 :(a) Long, narrow leaves are common in many monocots of intensely sunny areas. Shading is no problem, although the leaf bases probably do not photosynthesize at maximum efficiency. (b) These are the leaves of St. Augustine grass; the blades project horizontally and are attached to the stem by sheathing leaf bases. (c) In cattails, the sheathing leaf base can be half a meter long and provide firm but flexible attachment to the stem.

FIGURE 5: Maranta, a common houseplant, is a monocot that has broad leaves.
A leaf blade may be either simple or compound. A simple leaf has a blade of just one part (Fig. 6), whereas a compound leaf has a blade that is divided into several individual parts (Fig. 7).

FIGURE 6: Simple leaves have numerous shapes, some so deeply lobed as to be almost compound. Poplar (a) and oak (c) have one dominant midrib from which secondary vascular bundles extend. In the castor bean (b), the petiolar bundles immediately diverge into several main veins.
Think of how this might be affected by wind: If the blade is either large or flimsy, it twists and flexes and may tear. Tearing can be prevented by making the leaf small or tough or "pretorn" (compound). A compound leaf has many small blades, each attached by a petiolule to an extension of the petiole, the rachis. The combination of a petiolule and a small blade is a leaflet. Leaves may be palmately compound, with all leaflets attached at the same point, or pinnately compound, with leaflets attached individually along the rachis (Fig. 7).
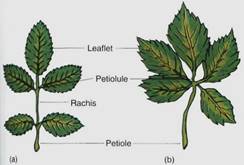
FIGURE 7: Leaflets of compound leaves have the same structure and metabolism as simple leaves. In pinnately compound leaves (rose, a) the petiolules of the leaflets are attached to the rachis, but in palmately compound leaves (Virginia creeper, b) they attach to the end of the petiole. In either type, any leaflet can be damaged or can abscise without affecting the remainder of the leaf.
Even a large compound leaf has only small leaflets, all of which can flex individually without tearing. Compound leaves have other advantages. When a mild breeze blows over two leaves of equal size, one simple and the other compound, it tends to flow smoothly over the simple leaf but turbulently over the more complex surface of the compound leaf. Turbulence brings in carbon dioxide and removes excess heat. Also, an insect can crawl or a fungus can spread across the entire blade of a simple leaf rather easily, but the edges and petiolule of a leaflet act as barriers that either prevent or at least slow movement to the rest of the leaf. In some plants, if pathogens severely damage a leaf, it abscises, carrying the pathogens with it and helping to keep pests away from healthy leaves.
Is it reasonable to conclude that plants with compound leaves are better adapted than plants with simple ones? Why are there any simple leaves at all? Most very large leaves are compound, apparently the only feasible architecture. The large simple leaves of banana are formed with a large, single lamina, but they have numerous special lines of weakness and are quickly torn to "compound" condition by wind. The largest simple leaves, those of the Victoria water lily, float on water and thus do not need large amounts of sclerenchyma for support, and adhesion to the water's surface prevents wind damage. Among medium and small leaves, simple ones are common, perhaps because a greater percentage of a simple leaf is composed of photosynthetic cells, whereas a compound leaf has a great deal of nonphotosynthetic rachis, petiolules, and edges.
Although most pinnately compound leaves are easily recognizable as leaves, some can be mistaken for a stem with simple leaves. Close examination reveals that leaflets never bear buds in the axils of their petiolules, and the tip of the rachis never has a terminal bud. Leaflets are always arranged in two rows, never in a spiral, whorled, or decussate phyllo-laxy.
Some of the tremendous range of shapes of leaves and leaflets is shown in Figure 8. There is speculation, but no total agreement, on how the different leaf shapes may be adaptive; many biologists believe that all function so well that none is clearly superior or strongly disadvantageous selectively. However, a large number of species have several type of leaves (Fig. 9a).
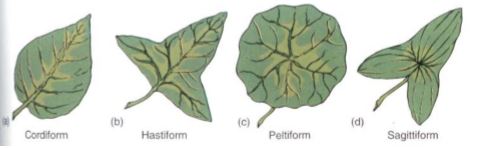
FIGURE 8: Four common leaf shapes; numerous other types also exist.
In the simplest cases, the first few leaves of a seedling are distinctly different from all leaves produced later; if they were merely smaller, it could be argued that the seedling does not have enough stored energy resources to construct the large leaves that a mature plant can afford. But very often the juvenile leaves differ from the adult leaves not only in size but also shape, texture, and even simple versus compound structure. It seems reasonable to hypothesize that juvenile leaves of seedlings are adaptive in the microhabitat close to the soil surface where a seedling is located, whereas adult leaves are adapted to the more aerial microhabitat inhabited by an older, larger plant.

FIGURE 9: (a) Beans have two types of leaves: the very first leaf formed by the seedling is simple (right), but all later leaves are compound (left). (b) This Azara lanceolata shoot has two types of leaves: Some leaves are large, whereas those closest to the axillary buds are small.
Other species produce two types of leaves simultaneously (Fig. 9b), often one type on long shoots and the other on short spur shoots. In cacti, the spines are short-shoot leaves, whereas the long-shoot leaves are green, fleshy, and in most species almost microscopic . In a small percentage of species, leaf shape and size are irregularly variable even on one stem, being influenced by environmental conditions that occur at the time of leaf initiation and expansion; amounts of sunlight and minerals are perhaps the most important factors.
Despite the numerous possibilities for variation within a species, leaf shape is a valuable tool for plant identification: The shape of the leaf, including its base and apex, are important, as is the nature of the margin. The margin may be entire (smooth), toothed, lobed, or otherwise modified (Fig. 10).
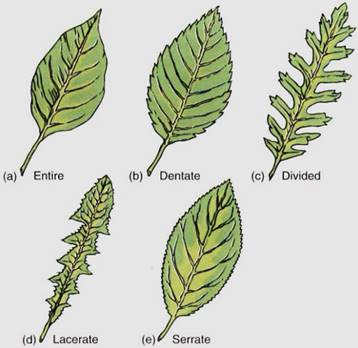
FIGURE 10:Several common types of leaf margins; in nature, hundreds of types exist, many being intermediate between two or among several other types.
Within a leaf are veins or bundles of vascular tissue; in a dicot they occur in a netted pattern called reticulate venation (Figs. 11 and 12). In monocots with long, strap-shaped leaves, the larger veins run side by side with few obvious interconnections: This is parallel venation (Figs. 13 and 14). Veins distribute water from the stem into the leaf and simultaneously collect sugars produced by photosynthesis and carry them to the stem for use or storage elsewhere.
At the leaf base, usually in the petiole, is an abscission zone oriented perpendicular to the petiole; its cells are involved in cutting off the leaf when its useful life is over (Fig. 15).

FIGURE 11:A leaf of Amaranthus retroflexus treated to remove all tissues except vascular bundles. The extensive reticulate venation is obvious in this dicot leaf; no part of the leaf is without veins. (Courtesy of D. Fisher, University of Hawaii)
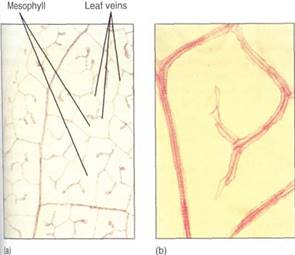
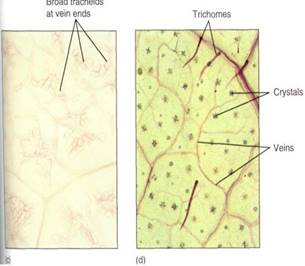
FIGURE 12:These leaves were treated (cleared) to make their cells transparent; then the xylem was stained to make the leaf veins visible. (a) In passionflower (Passiflora) leaves, the veins are narrow and highly branched. At this magnification (X 40), the height of the photograph represents 1 mm, so the veins occur close together. (b) Higher magnification, of privet leaf; at any point in the minor veins, only two or three tracheary elements are present (X 150). (c) These minor veins of crown of thorns (Euphorbia millii, a species that grows in deserts) are broad and have many wide tracheids at the ends (X 50). (d) This leaf of Acalypha has numerous crystals within the leaf and trichomes on the leaf surface (X 80).

FIGURE 13 : Cattails (Typha latifolia) have long, straplike leaves common in monocots. Vascular bundles run parallel to each other from leaf base to tip.
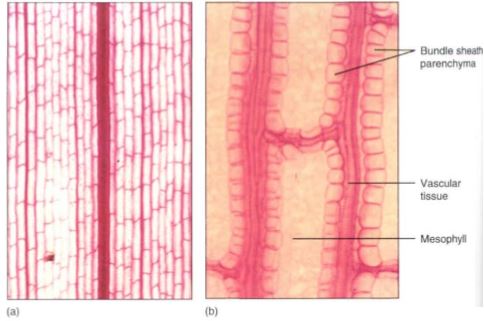
FIGURE 14: (a) Leaf clearing of corn (Zea), a monocot, with several sizes of longitudinal, parallel veins. If a longitudinal vein is broken by insect damage or any other problem, conduction can detour around the site by means of the fine transverse veins (X 50). (b) Higher magnification showing the parenchyma cells that surround the leaf bundles in corn (X 250).
In autumn, as deciduous leaves begin to die, abscission zone cells release enzymes that weaken their walls. As the leaf twists in the wind, cell walls break and the leaf falls of Adjacent undamaged cells swell and become corky, forming protective scar tissue, the leaf scar, across the wound . Without an abscission zone, dead leaves might tear off irregularly, leaving an open wound vulnerable to pathogens.
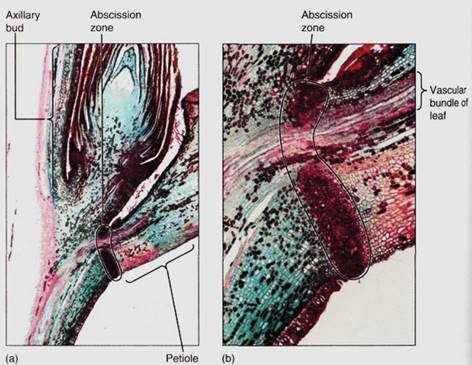
FIGURE 15 :(a) Longitudinal section through a leaf axil, showing the abscission zone in the petiole and the axillary bud just above (X 15). (b) Magnification of the abscission zone while the leaf is still healthy. The xylem and phloem are not interrupted until the leaf actually falls away (X 80).



|
|
|
|
للعاملين في الليل.. حيلة صحية تجنبكم خطر هذا النوع من العمل
|
|
|
|
|
|
|
"ناسا" تحتفي برائد الفضاء السوفياتي يوري غاغارين
|
|
|
|
|
|
|
بمناسبة مرور 40 يومًا على رحيله الهيأة العليا لإحياء التراث تعقد ندوة ثقافية لاستذكار العلامة المحقق السيد محمد رضا الجلالي
|
|
|