Levels of Taxonomic Categories
Plants have varying degrees of relatedness, and a natural classification system reflects this in its numerous levels.
The most fundamental level of classification is the species, which ideally and theoretically is a set of individuals closely related by descent from a common ancestor. Members of a species can interbreed with each other successfully but cannot interbreed with individuals of any other species (Table 18.1). Most species are not so predictable; they may not interbreed well with closely related species, but an occasional cross-pollination results in a viable seed that grows into a fertile adult. If this occurs frequently, the two plant groups may best be considered subspecies of a single species. The two subspecies must be so similar genetically that the chromosomes of the two parents can function in the same nucleus and, when the plant is mature, spore mother cells can undergo meiosis successfully. For all this to happen, the organisms must be extremely closely related, having undergone divergent evolution from a common ancestor only recently, so that few mutations have accumulated since they diverged. But as mutations continue to accumulate in each subspecies, finally either no viable seed results when the two are crossed or the seed glows into a sterile plant. The two then must be considered separate species.
Closely related species are grouped together into genera (singular: genus; Table 18.1). Deciding whether several species are closely related enough to be placed together in the same genus is difficult.
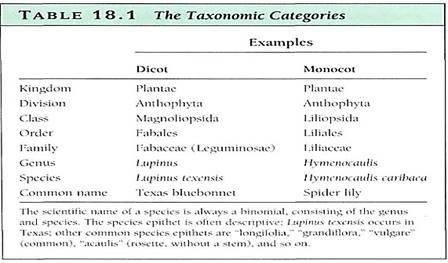
No objective criteria exist; the decision is entirely subjective and often the cause of great dispute. Some taxonomists, generally referred to as "lumpes" believe that even relatively distantly related species should be grouped together in large genera. Other taxonomists, called "splitters," prefer to have many small genera, each containing only a few species that are extremely closely related. For example, some taxonomists believe that cranberries and blueberries are so similar that they should go into the same genus, Vaccinium; others think that cranberries are distinct enough that segregating them into their own genus, Oxycoccus, more accurately reflects evolutionary reality. Both groups of taxonomists agree that the two sets of species are closely related, but they have different opinions as to how much evolution has occurred since the time of the most recent common ancestor. The critical concern is that the genera are natural, that all the species included in the genus are related to each other by a common ancestor, and that all descendants of that common ancestor are in the same genus. Such a group is monophylelic (Fig. 1). In an unnatural, polyphyletic group, members have evolved from different ancestors and may resemble each other only as a result of convergent evolution.
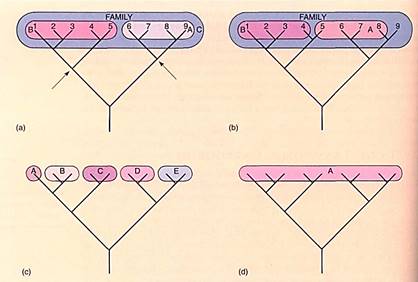
FIGURE 1: (a) In this diagram two natural genera, A and B, have been designated. Both are natural because all members of each can be traced back to common ancestors (arrows). The two genera would constitute a family, C, which would also be natural. (b) Genus A is not natural because it is not monophyletic: species 5 does not share an appropriate common ancestor with the others; tee is a common ancestor, but it is in the remote past, at the family level. Technically, it is acceptable to leave species 9 out, making it a monospecific genus (a genus with only one species), but because it is even more closely related to species 8 than are 6 and 7, leaving it out would be inconsistent Genus B is monophyletic, so it is a natural group, but species 5 should be included for consistency. Even though the genera are either incomplete or polyphyletic, still the family, C, is perfectly monophyletic and complete. (c) The family classified by a "splitter." The concept of a monospecific genus is valid: If genera B and C were extinct, then A should be in a genus by itself because it is not closely related to any other living species, (d) The family classified by an extreme "lumper"; this is also a valuable concept. If rapid adaptive radiation is occurring, then all species may be so similar that no strong distinctions exist to use as guidelines for creating smaller genera.
The level above genus is family, each family being composed of one, several, or often many genera. Most families are well defined, with widespread agreement as to which species and genera belong in a particular family. As examples, consider how easy it is to recognize the following families: cacti, orchids, daisies, palms, and grasses. The reason for this probably is the age of most families. A line of evolution must be very old—usually at least several tens of millions of years—to diversify into several genera and many species. Even closely related families have been separate evolutionary lines for at least 20 or 30 million and often 50 million years. The common ancestor and many of the earliest species probably have become extinct. Therefore, although two families have many characters in common, enough features are unique to each family to make them recognizable as distinct from each other (Fig. 2). Problems arise when the earliest species and genera of a family persist; these have characteristics intermediate between the families, so it can be argued reasonably that all should be considered a single family. The term "subfamily" is used for the groups of closely related genera within such a single large family.
The levels above family are order, class, division, and kingdom. Although you might expect universal agreement at the level of kingdom, even the boundaries of the plant kingdom, kingdom Plantae, are disputed. Some believe that the green algae should be included because they are almost identical biochemically to vascular plants. Others believe that, despite the biochemical similarity, the green algae should be excluded because they are so different morphologically and anatomically.
The classification used in this book is based on that proposed by Drs. H. Bold, C. Alexopoulos, and T. Delevoryas in Morphology of Plants and Fungi (1987), which recognizes a large number of divisions. Other botanists have proposed systems with many fewer divisions, with many of the divisions of Bold, Alexopoulos, and Delevoryas considered subdivisions. There is no "correct" view; it is most important to understand the reasons used to support each view.
Except for kingdom, genus, and species, the names must have a certain ending to indicate the classification level. Division names end in -phyla, for example Chlorophyta (green algae), Coniferophyta (conifers), and Anthophyta (flowering plants) (Table 18.1). Class names end in -opsida, subclass names in -idae, order names in -ales, and family names in -aceae (pronounced as if you were spelling the word "ace"). Genera and species names do not have standard endings. The scientific name of a species is its genus and species designations used together and either underlined or italicized; for example, tomato is Lycopersicon escuientum. Note that the species name is not "esculentum" but Lycopersicon esculentum. Esculentum is the species epithet, the word that distinguishes this species only from the other species of the genus Lycopersicon. We cannot refer to tomatoes as "esculentum" because that species epithet is used for many different species in different genera: for example, buckwheat—Fagopyrum esculentum, and taro—Colocasia esculenta.
Some families have two names; the above rules were adopted to regularize family names so that each family is named after one of its genera, using the -aceae suffix. The rose family was named Rosaceae, based on the genus Rosa. This worked well for most families, but for some, the old name was so well-known and so familiar that it was kept as well. Examples are Asteraceae (Compositae), Fabaceae (Leguminosae, legumes), Aracaceae (Palmae, palms), Poaceae (Gramineae, grasses), Brassicaceae (Cruciferae, mustards), and Apiaceae (Umbelliferae, umbels).
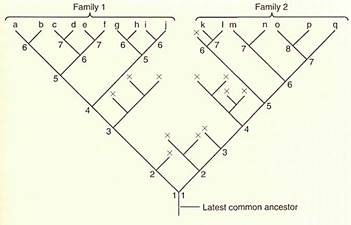
FIGURE 2: In this hypolhetical evolutionary history, two lines diverged long ago, then many of the early species became extinct (lines marked with x), and only some continued on, speciating into two separate families. In family 1 all extant species have at least six speciation events in their history since the latest common ancestor shared with family 2. Family 2 has undergone slightly more rapid evolution: The minimum number of branch points is seven, and two species have eight. Thus, since these two lines of evolution diverged, many changes have occurred in each, so they are quite distinct. In reality, there are about 400,000 species of vascular plants with unknown hundreds of thousands or millions of extinct species, so the true phylogeny is gigantic. The latest common ancestor for vascular plants probably occurred about 420 million years ago.
 الاكثر قراءة في مواضيع عامة في علم النبات
الاكثر قراءة في مواضيع عامة في علم النبات
 اخر الاخبار
اخر الاخبار
اخبار العتبة العباسية المقدسة





















