Adriamycin
1. Introduction
Adriamycin is the trade name for an anthraquinone-containing antibiotic which also has the international nonproprietary name (INN) of doxorubicin (1) and the NCI internal identification number NSC 123127. It is a member of the anthracycline group of compounds that contain an anthraquinone chromophore and a polycyclic ring system (1). It has good anticancer activity against a wide spectrum of tumors (and still remains as the antitumor agent that has the widest spectrum of anticancer activity) and is one of the most extensively used of the fifty or so chemotherapeutic compounds currently in clinical use (2-4). Its full potential as an anticancer agent has not been reached because of a dose-limiting cardiotoxicity (1-4). Despite enormous effort during the last 30 years to develop more effective and less cardiotoxic derivatives, it remains one of the best, proven anticancer drugs. It also exhibits good antibacterial activity.
2. Structure and Chemistry
Adriamycin consists of a planar tetracyclic ring system linked by a glycosidic bond to daunosamine, an amino sugar (Fig. 1). It is a bright red compound that is normally isolated as the hydrochloride salt, has the chemical composition C27H29O11·HCl, a molecular weight of 579.98, and is soluble in water, physiological saline, and methanol (5, 6). The pKa of the amino group is 8.2 (5, 6). In neutral aqueous buffers, it self-associates (7), has absorbance maxima at 233, 480, and 530 nm, and at concentrations less than 25 µM has an extinction coefficient of 11,500 M–1cm–1at 480 nm (7). It is fluorescent and can be quantitated in aqueous solutions using excitation and emission wavelengths of 480 and 595 nm, respectively. The solid compound is quite stable if kept in the dark at 4°C and free of moisture. It is photosensitive and therefore solutions must also be kept in the dark at 4°C; even under these conditions, some decomposition will occur in a week or so. It chelates Fe3+with exceedingly high affinity and readily forms the Fe(Adriamycin)3 complex (8).

Figure 1. Structure of Adriamycin and clinically relevant derivatives.
3. History
In 1958 a new species of Streptomyces was identified in a soil sample taken from southern Italy. A red antibiotic, daunomycin was isolated from this microorganism and exhibited good activity against a range of murine tumors (1). In a search for other potentially more active derivatives of daunomycin, the soil microorganism was subjected to the mutagen N-nitroso-N-methyl urethane and produced a mutant strain that yielded a modified form of the antibiotic. This new antibiotic (named Adriamycin because of the closeness of the original soil sample to the Adriatic Sea) exhibited both a wider spectrum of activity and an improved anticancer response against animal tumors than daunomycin (1) and was also remarkably successful in treating human tumors. Adriamycin was rapidly introduced into clinical trials in Italy and the United States and was approved for clinical use in the United States in 1974 (1).
4. Clinical Use
4.1. Tumors
Adriamycin is particularly useful for treating solid tumors such as breast, lung, ovarian, and thyroid carcinomas, as well as soft tissue sarcomas (2-4). It is also active against lymphoid and myelogenous leukemia (2-4).
4.2. Dose and Administration
It is normally administered as a bolus at a dosage of 45–75 mg/m2 every 3–4 weeks, or more recently as a continuous intravenous infusion during 4–5 days (2, 3). It is inactive if administered orally because the glycosidic bond is hydrolyzed in the gastrointestinal tract. For most treatment regimes it is administered as one component of a combination of drugs (2).
4.3. Side Effects
The most significant problem is a cumulative, dose-dependent cardiomyopathy that can lead to heart failure in up to 10% of patients who receive the maximum recommended dose of 550 mg/m2(2, 3). This cardiotoxicity can be acute (hours to days following treatment) or more commonly, can be delayed by months or even years. The cardiotoxicity appears to be due to the redox activity of the drug (9): Adriamycin can undergo a one electron reduction [catalyzed by a range of enzymes such as microsomal cytochrome P450 reductase, mitochondrial NADH dehydrogenase, cytochrome b5 reductase, and xanthine reductase (10)] leading to production of the semiquinone that generates superoxide in the presence of molecular oxygen, then hydrogen peroxide, and ultimately the extremely reactive and highly toxic hydroxyl radical. These hydroxyl radicals react with any nearby molecule, and hence lead to oxidative damage of critical targets such as DNA and membrane lipids. In most tissues this lethal process is minimized by superoxide dismutase (which converts superoxide to hydrogen peroxide) and catalase (which subsequently converts hydrogen peroxide to water), as well as glutathione peroxidase (which also converts hydrogen peroxide to water). Heart tissues are particularly sensitive to Adriamycin because of their greatly compromised capacity for protection against hydroxyl radicals due to the lack of the bulk hydrogen peroxide detoxifying enzyme catalase and also because Adriamycin inhibits gutathione peroxidase activity (10). Other side effects of Adriamycin therapy are hair loss (alopecia) which usually starts after the first dose of Adriamycin, nausea and vomiting (usually overcome nowadays by appropriate antinausea drugs), and reduction in bone marrow function (with maximum suppression 10–14 days following treatment).
4.4. Pharmacokinetics
The disappearance of Adriamycin in plasma is adequately described by a three-compartment model whose half-lives are approximately 10 min, 3 h, and 30 h (11). The plasma level is rapidly depleted and only 0.2 mM remains (average of nine patients) several hours after administration of a 30 mg/m2 bolus dose of Adriamycin (12); 0.02 µM remains several hours following a 75 mg dose (either as a bolus or 4 h infusion) to a single patient (13). In contrast, the intracellular concentration of drug was approximately 4 µM and 6 µM, respectively, for these two studies (12, 13). The striking features of these results are that the intracellular concentration of drug is typically 100–1000 times greater than that in plasma (14), and that the intracellular drug pool has an extremely long half-life of 4–5 days (13). The uptake into solid tumors is similar to that of normal tissues (12).
Pharmacology. The major metabolite of Adriamycin involves metabolism of the C9 side chain alcohol to adriamycinol, where the conversion is catalyzed by the ubiquitous cytoplasmic NADPH-dependent aldo-keto reductase (15). Subsequent microsomal glycosidases that are present in most tissues convert adriamycinol into the inactive deoxyadriamycinol aglycon and daunosamine. The aglycon is then demethylated and conjugated to polar groups to yield more hydrophilic metabolites that are excreted mainly in the bile (15).
The drug distribution has been examined in 20 cancer patients where tissue uptake decreased in the following order: liver > lymph nodes > muscle and bone marrow > fat and skin (12). It has long been known that Adriamycin localizes in the nucleus at the subcellular level (16). This has been confirmed recently in single squamous carcinoma cells by quantitative confocal laser scanning microscopy, where the nuclear level was twice that of the cytoplasm 30 minutes after exposure to 1 µg/mL of Adriamycin, and increased even more with time (17). Interestingly, the cytoplasmic drug level was also higher than that of the medium (17), and this is surprising because Adriamycin is taken up into cells by passive diffusion (18). Nuclear localization is readily attributed to the known high affinity of Adriamycin for DNA (see below). The nature of cytoplasmic localization is less clear, but the distribution as a multitude of small patches of fluorescence throughout the cytoplasm (19) is consistent with some degree of mitochondrial localization (because the drug binds to mitochondrial membranes and, to a lesser extent, also to mitochondrial DNA); it is known that there are as many as a thousand mitochondria per cell (20).
4.5. Resistance
When cells in culture are exposed to Adriamycin for an extended period of time, the cells gradually become resistant to the drug. This type of resistance is due primarily to amplification of the multidrug transporter gene mdr1 that leads to overexpression of a 170-kDa glycoprotein now known as P-glycoprotein (21). This protein forms a pore in the cell membrane and actively pumps Adriamycin (and many other drugs) out of the cell, using ATP as an energy source. In patients treated with Adriamycin, much of the resistance that develops over several months (and diminishes the antitumor effect of the drug) and appears to derive from overexpression of P-glycoprotein. To enhance the usefulness of Adriamycin, there have been some attempts to develop specific inhibitors of this protein. This approach is no longer considered viable because of the multitude of other drug efflux pumps that have been identified in recent times (22).
In addition to the phenomenon of multidrug resistance, other mechanisms of resistance have been identified for Adriamycin, including elevated levels of glutathione and decreased levels of topoisomerase II in cells in culture (although not yet demonstrated in tumors) (21).
5. Interactions with DNA
Numerous studies have shown that Adriamycin binds to DNA with high affinity by intercalating between adjacent base pairs of DNA. In this process the DNA unwinds to accommodate the drug. This is reflected by an increase in the length of small linear fragments of DNA (hence an increase in the viscosity of the solution) or in the loss of a number of negative supercoils for supercoiled DNA such as a plasmid (that results in an initial increase of viscosity and a decrease of the sedimentation coefficient). These processes have been summarized previously (3, 23). The structure of the intercalated species has been fully characterized by X-ray crystal studies of the drug–oligonucleotide complex. The dominant features are that the drug chromophore lies virtually at right angles to the adjoining base pairs and the amino sugar fits snugly into the minor groove of DNA (24, 25).
Earlier binding studies (before about 1980) misinterpreted curved Scatchard plots as indicating more than one class of DNA binding site. It is now known that this curvature which results from the extreme form of negative cooperativity is displayed when the drug binds to DNA, and is fully described by the neighbor-exclusion principle; values for the intrinsic association constant and the number of occluded base pairs are approximately 2·106 M–1and 3.0 bp, respectively, at near physiological ionic strengths (26, 27). Although this interpretation assumes that the drug intercalates randomly between all possible base pair combinations, this is not strictly correct because there is a slight selectivity of sequence for some sites. Theoretical quantum mechanical calculations indicated the requirement for a three base pair site, and ACG is likely to be the preferred site (28, 29). In vitro transcription footprinting studies confirmed the requirement for a triplet site, but showed that the preferred consensus sequence is TCA (where the drug intercalates between C and A) (30), although there was clearly only small energetic differences between this and other triplet sites.
Adriamycin is in rapid equilibrium with DNA. The on-rate is diffusion controlled, and the off-rate is also fast and usually measured by stopped-flow detergent sequestration. This has revealed that Adriamycin has an overall half-life on DNA of approximately 0.5 s at room temperature (31), although by analogy with the structurally similar drug daunomycin, the kinetic processes are likely to be much more complex, especially for longer dissociation times (32(.
6. Mechanism of Action
The exact molecular events involved in the mechanism of action of Adriamycin have not yet been resolved. Although many possible mechanisms have been proposed and the problems in identifying the critical factors have been well summarized (33, 34), many potential mechanisms have been identified, including impairment of topoisomerase II activity, bioreductive action of the drug, free radical effects, membrane related effects (33, 34). There is a convincing body of evidence to show that the primary target is DNA (33-35). Some of this evidence is that almost all of the drug in the nucleus (>99.8% of single, living cells is associated with DNA (36); more than 80% of all of the drug present in human tumor biopsies is associated with DNA (37); and increasing drug activity) over three orders of magnitude) correlates with increasing DNA binding, damage, or impairment of DNA template activity (38). Therefore DNA binding appears to be related in some way to the anticancer properties, and at present there are two types of interactions with DNA that appear to relate to the drug action: impairment of topoisomerase II activity and formation of drug–DNA adducts.
6.1. Impairment of Topoisomerase II
When Adriamycin is added to tumor cells in culture, protein-associated double- and single-strand DNA breaks occur close to nuclear matrix attachment sites where topoisomerase II is localized. This enzyme regulates the topological state of DNA, and the DNA breaks appear to arise from intercalation of the drug at that site, which results in structural distortions that interfere with the re-ligation step of topoisomerase II (2, 39, 40). There are several reasons why this appears to be a major contribution to the mechanism of action of Adriamycin: the activity of the enzyme is reduced in tumor cells which are resistant to Adriamycin (39, 40); a correlation has been shown between the induction of double-strand DNA breaks and the cytotoxicity of Adriamycin in P388 leukemia cells (41); and this type of DNA damage occurs at clinical levels of the drug (39, 40). However, there is also evidence that other factors contribute to the mechanism of action of Adriamycin and that the activity cannot be accounted for solely by impairment of topoisomerase II: some studies failed to detect DNA strand breaks in cell cultures at Adriamycin concentrations where cytotoxic responses were observed (42); topoisomerase II mediated damage is rapidly reversible following removal of the drug (39), whereas it is known that DNA double- strand breaks increase long after removal of the drug (43); some derivatives that induce a high level of double-strand breaks exhibit a low level of cytotoxicity (44); and there is little evidence that topoisomerase II is involved in some tumors (10).
6.2. Adriamycin–DNA Adducts
There have been many reports of Adriamycin–DNA adducts formed by enzymatic, microsomal, or cellular activation of the drug (33, 45, 46), but it was not until 1990 that it became clear from in vitro transcription footprinting studies that these adducts formed predominantly at 5′-GC-3′ sequences (47). It was subsequently shown that adducts at these sites behave as an interstrand cross-link (48,( 49, although with limited stability [half-life of 5 –40 h depending on the DNA sequence and fragment length (49, 50)]. It is now clear that adducts at GC sequences and interstrand cross-links at GC sequences are one and the same lesion and that the adducts stabilize DNA sufficiently so that they function as “virtual interstrand cross-links” (51-53). The adducts have been well characterized in the solid state by X-ray crystal diffraction (54) and in solution by 2-D NMR (55).
Surprisingly, it has been shown that the adducts are mediated by formaldehyde and involve a characteristically unstable aminal linkage to the N-2 of guanine on one strand of DNA (51-55) (Fig. 2). This unusual adduct exhibits some of the characteristics of an interstrand cross-link because of the additional stability that arises from the intercalated chromophore and from additional hydrogen bonds that are formed to the second DNA strand (51-55). The cross-linkage is unstable to heat and to alkali, and this explains why it has proven so difficult to find this lesion in cells in the past (50). Conditions have been established to isolate these lesions from tumor cells (56-59), and this led to a resurgence in cellular studies of this drug. There is now good evidence that these adducts contribute to the mechanism of action of Adriamycin: the capacity of different derivatives of Adriamycin to form adducts (virtual cross-links) correlates well with their cytotoxicity (57), and gene-specific cross-linking assays have detected sufficient lesions in both the nuclear and mitochondrial genomes to show that this lesion is cytotoxic at clinical levels of the drug (58). Total cellular DNA adducts have also been quantitated directly using [14C]Adriamycin (58). Because of the formaldehyde-dependent activation of Adriamycin, formaldehyde-releasing prodrugs that have been examined in combination with Adriamycin lead to as much as a 20-fold increase of Adriamycin–DNA adduct levels and a synergistic cytotoxic response (59).
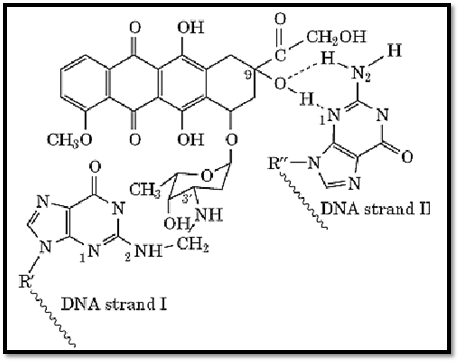
6.3. Other Possible DNA-Related Effects
Two additional DNA-related effects also occur at low drug levels in vitro, but at this stage the contribution to the mechanism of action of Adriamycin under clinical conditions is unknown (34): impairment of DNA ligase activity (60): impairment of helicase activity (61).
6.4. Current Status
The evidence at present indicates that there is no single mechanism of action of Adriamycin. Both impairment of topoisomerase II activity (resulting in protein associated double-strand breaks) and formation of interstrand cross-links occur at clinical levels of the drug. It is known that Adriamycin activates the general cellular response to DNA damage in which the tumor suppressor protein p53 is induced, resulting in arrest of the cell cycle and the induction of apoptosis (62). Although it is known that apoptosis occurs at submicromolar concentrations of the drug (63), this response is yet to be fully understood in the context of drug-induced cytotoxicity (64). The development of new derivatives of Adriamycin in the future is likely to rely heavily on an even more detailed understanding of the molecular consequences of the DNA damage induced by this drug.
7. The Search for New Derivatives
Because of the dose-limiting toxicity of Adriamycin, there has been an intense international effort in the past three decades to find new derivatives that have improved anticancer activity and/or reduced cardiotoxicity. This endeavor has been extremely well summarized by Weiss (1). Unfortunately, the outcome from this search has been disappointing. From more than 2000 derivatives tested by 1992 (and perhaps as many as 2500 tested to date), none is substantially superior to Adriamycin for proven clinical anticancer activity. However, several offer advantages for clinical use: idarubicin (lacking the methoxy group at the C4 position) is more amenable to oral administration than Adriamycin; epirubicin is less cardiotoxic and results in less nausea and vomiting than Adriamycin at equimolar doses—it has anticancer activity similar to that of Adriamycin, but reduced side effects have facilitated its wide use as an alternative to Adriamycin. High-dose clinical trials are still in progress with this derivative. One new and encouraging approach that is emerging is the development of “preactivated” forms of the anthracyclines; these formaldehyde-activated derivatives (doxoform, epidoxoform) exhibit increased toxicity to tumor cells, especially to anthracycline-resistant cells (65).
References
1. R. B. Weiss (1992) Semin. Oncol. 19, 670–686.
2. V. T. DeVita, S. Hellman, and S. A. Rosenberg (2001) Cancer: Principles and Practice of Oncology, 6 ed., Lippincott Williams and Wilkins, Philadelphia.
3. W. B. Pratt, R. W. Ruddon, W. D. Ensminger, and J. Maybaum (1994) The Anticancer Drugs, 2 ed, Oxford University Press, New York, pp. 155–165.
4. T. W. Sweatman and M. Israel (1997) In Cancer Therapeutics, Experimental and Clinical Agents (B.A. Teicher, ed.), Humana Press, Totowa, NJ, pp. 113–135.
5. F. Arcamone (1982) Doxorubicin: Anticancer Antibiotics, Academic Press.
6. R. T. Dorr and D. S. Alberts (1982) In Current Concepts in the Use of Doxorubicin Chemotherapy (S.E. Jones, ed.), Farmitalia Carla Erba S.p.A., Milano, Italy, pp. 1–20.
7. J. B. Chaires, N. Dattagupta, and D. M. Crothers (1982) Biochemistry 21, 3927–3932.
8 A. Garnier-Suillerot (1988) In Anthracycline and Anthracenedione-Based Anticancer Agents (J.W. Lown, ed.), Elsevier, Amsterdam, pp. 129–161.
9. J. H. Doroshow (1995) In Anthracycline Antibiotics: New Analogues, Methods of Delivery, and Mechanisms of Action (W. Priebe, ed.), American Chemical Society, Washington, DC, pp. 259267- .
10. pp. 160–164 of Ref. 3.
11. p. 378 of Ref. 2.
12. Y. N. Lee, K. K. Chan, P. A. Harris, and J. L. Cohen (1980) Cancer 45, 2231–2239.
13. P. A. J. Speth, P. C. M. Linssen, J. B. M. Boezeman, J. M. C. Wessels, and C. Haanen (1986)377, . 414–422
14. C. E. Myers, E. G. Mimnaugh, G. C. Yeh, and B. K. Sinha (1988) In Anthracycline and Anthracenedione-Based Anticancer Agents (J.W. Lown, ed.), Elsevier, Amsterdam, p. 529.
15. pp. 158–160 of Ref. 3.
16. A. DiMarco (1975) Cancer Chemother. Rep. 6, 91–106.
17. K. Kawai, Y. Minamiya, M. Kitamura, I. Matsuzaki, M. Hashimoto, H. Suzuki, and S. Abo (1997) Cancer 79, 214–219.
18. M. Dalmark and H. H. Storm (1981) Gen. Physiol. 78, 349–364.
19. V. Pillay, R. D. Martinus, J. S. Hill, and D. R. Phillips (1998) J. Cell. Biochem. 69, 1–7.
20. J. W. Shay and H. Werbin (1987) Mutat. Res. 186, 149–160.
21. pp. 50–66 of Ref. 3.
22. J. S. Lee, S. Scala, Y. Matsumoto, B. Dickstein, R. Robey, Z. Zhan, G. Altenberg, and S. E. Bates (1997) J. Cell Biochem. 513–526.
23. S. Neidle and M. R. Sanderson (1983) In Molecular Aspects of Anti-Cancer Drug Action (S. Neidle and M.J. Waring, eds.), Macmillan, London, pp. 35–56.
24. A. H. Wang, G. Ughetto, G. J. Quigley, and A. Rich (1987) Biochemistry 26, 1152–1163.
25. G. Ughetto (1988) In Anthracycline and Anthracenedione-Based Anticancer Agents (J.W. Lown, ed.), Elsevier, Amsterdam, pp. 295–334.
26. F. Barcelo, J. Martorell, F. Gavilanes, and J. M. Gonzalez-Ros (1988) Biochem. Pharmacol. 372133–2138 , .
27. E. Stutter, H. Schuetz, and H. Berg (1988) In Anthracycline and Anthracenedione-Based Anticancer Agents (J.W. Lown, ed.), Elsevier, Amsterdam, pp. 245–293.
28. K. Chen, N. Gresh, and B. Pullman (1986) Nucleic Acids Res. 14, 2251–2267.
29. B. Pullman (1991) Anti-Cancer Drug Design 7, 95–105.
30. H. Trist and D. R. Phillips (1989) Nucleic Acids Res. 17, 3673–3688.
31. B. Gandecha and J. R. Brown (1985) Biochem. Pharmacol. 34, 733–736.
32. D. R. Phillips, P. Greif, and R. C. Boston (1988) Mol. Pharmacol. 33, 225–230.
33. pp. 528–569 of Ref. 14.
34. D. A. Gewirtz (1999) Biochem. Pharmacol. 57, 727–741.
35. C. Cullinane, A. van Rosmalen, and D. R. Phillips (1994) Biochemistry 33, 4632–4638.
36. M. Gigli, S. M. Doglia, J. M. Millot, L. Valantini, and M. Manfait (1988) Biochim. Biophys. Acta 950, 13–20.
37. J. Cummings and C. S. McArdle (1986) Br. J. Cancer 53, 835–838.
38. L. Valentini, V. Nicolella, E. Vannini, M. Menozzi, S. Penco, and F. Arcamone (1985) Il. Farmaco Ed. Sci. 40, 377–389.
39. C. Holm, J. Covey, D. Kerrigan, K. W. Kohn, and Y. Pommier (1991) In DNA Topoisomerases in Cancer (M. Pomesil and K. Kohn, eds.), Oxford University Press, pp. 161–171.
40. Y. Pommier (1995) In Anthracycline Antibiotics: New Analogues, Methods of Delivery and Mechanisms of Action. ACS Symposium Series No. 574, pp. 183–203.
41. G. J. Goldenberg, H. Wang, and G. W. Blair (1986) Cancer Res. 46, 2978–2983.
42. F. A. Fornari, W. D. Jarvis. M. S. Orr, J. K. Randolph, S. Grant, and D. A. Gerwitz (1996) Biochem. Pharmacol. 51, 931–940.
43. M. Binaschi, G. Capranico, P. De Isabella, M. Marini, R. Supino, and S. Tinelli (1990) Int. J. Cancer 45, 347–352.
44. M. Binaschi, G. Capranico, L. Dal Bo, and F. Zunino (1997) Mol. Pharmacol. 51, 1053–1059.
45. D. R. Phillips (1990) In Molecular Basis of Specificity in Nucleic Acid-Drug Interactions (B. Pullman and J. Jortner, eds.), Kluwer Academic, Dordrecht, The Netherlands, pp. 137–155.
46. J. Cummings, L. Anderson, N. Willmott, and J. F. Smyth (1991) Eur. J. Cancer 27, 532–535.
47. C. Cullinane and D. R. Phillips (1990) Biochemistry 29, 5638–5646.
48. C. Cullinane, A. van Rosmalen, and D. R. Phillips (1994) Biochemistry 33, 4632–4638.
49. S. M. Cutts and D. R. Phillips (1995) Nucleic Acids Res. 23, 2450–2456.
50. A. van Rosmalen, C. Cullinane, S. M. Cutts, and D. R. Phillips (1995) Nucleic Acids Res. 23 42–50, .
51. D. J. Taatjes, G. Guadiano, K. Resing, and T. H. Koch (1996) J. Med. Chem. 39, 4135–4138.
52. D. J. Taatjes, G. Guadiano, K. Resing, and T. H. Koch (1997) J. Med. Chem. 40, 1276–1286.
53. R. A. Luce, S. Th. Sigurdsson, and P. B. Hopkins (1999) Biochemistry 38, 8682–8690.
54. A. H. Wang, Y. Gao, Y. Liaw, and Y. Li (1991) Biochemistry 30, 3812–3815.
55. S. M. Zeman, D. R. Phillips, and D. M. Crothers (1998) Proc. Natl. Acad. Sci. USA 95, 11561 . 11565-
56. A. Skladanowski and J. Konopa (1994) Biochem. Pharmacol. 47, 2269–2278.
57. A. Skladanowski and J. Konopa (1994) Biochem. Pharmacol. 47, 2279–2287.
58. C. Cullinane, S. M. Cutts, C. Panousis, and D. R. Phillips (2000) Nucleic Acids Res. 28, 1019.1025-
59. S. M. Cutts, A. Rephaeli, A. Nudelman, I. Hmelnitsky, and D. R. Phillips (2001) Cancer Res., in press.
60. G. Ciarrochi, M. Lestingi, M. Fontana, S. Spadari, and A. Montecucco (1992) Biochem J. 279 141–146- , .
61. N. R. Bachur, R. Johnson, F. Yu, R. Hickey, N. Appelgren, and L. Malkas (1993) Mol. Pharmacol. 44, 1064–1069.
62. G. Zaleskis, E. Berleth, S. Verstovek, M. J. Ehrke, and E. Mihich (1994) Mol. Pharmacol. 46 901–908, .
63. A. Skladanowski and J. Konopa (1993) Biochem. Pharmacol. 46, 375–382.
64. R. B. Lock and L. Stribinskiene (1996) Cancer Res. 56, 4006–4012.
65. D. J. Taatjes and T. H. Koch (2001) Curr. Med. Chem. 8, 15–29.





















