























النبات

مواضيع عامة في علم النبات
الجذور - السيقان - الأوراق
النباتات الوعائية واللاوعائية
البذور (مغطاة البذور - عاريات البذور)
الطحالب
النباتات الطبية
الحيوان
مواضيع عامة في علم الحيوان
علم التشريح
التنوع الإحيائي
البايلوجيا الخلوية
الأحياء المجهرية
البكتيريا
الفطريات
الطفيليات
الفايروسات
علم الأمراض
الاورام
الامراض الوراثية
الامراض المناعية
الامراض المدارية
اضطرابات الدورة الدموية
مواضيع عامة في علم الامراض
الحشرات
التقانة الإحيائية
مواضيع عامة في التقانة الإحيائية
التقنية الحيوية المكروبية
التقنية الحيوية والميكروبات
الفعاليات الحيوية
وراثة الاحياء المجهرية
تصنيف الاحياء المجهرية
الاحياء المجهرية في الطبيعة
أيض الاجهاد
التقنية الحيوية والبيئة
التقنية الحيوية والطب
التقنية الحيوية والزراعة
التقنية الحيوية والصناعة
التقنية الحيوية والطاقة
البحار والطحالب الصغيرة
عزل البروتين
هندسة الجينات
التقنية الحياتية النانوية
مفاهيم التقنية الحيوية النانوية
التراكيب النانوية والمجاهر المستخدمة في رؤيتها
تصنيع وتخليق المواد النانوية
تطبيقات التقنية النانوية والحيوية النانوية
الرقائق والمتحسسات الحيوية
المصفوفات المجهرية وحاسوب الدنا
اللقاحات
البيئة والتلوث
علم الأجنة
اعضاء التكاثر وتشكل الاعراس
الاخصاب
التشطر
العصيبة وتشكل الجسيدات
تشكل اللواحق الجنينية
تكون المعيدة وظهور الطبقات الجنينية
مقدمة لعلم الاجنة
الأحياء الجزيئي
مواضيع عامة في الاحياء الجزيئي
علم وظائف الأعضاء
الغدد
مواضيع عامة في الغدد
الغدد الصم و هرموناتها
الجسم تحت السريري
الغدة النخامية
الغدة الكظرية
الغدة التناسلية
الغدة الدرقية والجار الدرقية
الغدة البنكرياسية
الغدة الصنوبرية
مواضيع عامة في علم وظائف الاعضاء
الخلية الحيوانية
الجهاز العصبي
أعضاء الحس
الجهاز العضلي
السوائل الجسمية
الجهاز الدوري والليمف
الجهاز التنفسي
الجهاز الهضمي
الجهاز البولي
المضادات الميكروبية
مواضيع عامة في المضادات الميكروبية
مضادات البكتيريا
مضادات الفطريات
مضادات الطفيليات
مضادات الفايروسات
علم الخلية
الوراثة
الأحياء العامة
المناعة
التحليلات المرضية
الكيمياء الحيوية
مواضيع متنوعة أخرى
الانزيمات

Glycogenesis and Glycogenolysis Regulation
 المؤلف:
Denise R. Ferrier
المؤلف:
Denise R. Ferrier
 المصدر:
Lippincott Illustrated Reviews: Biochemistry
المصدر:
Lippincott Illustrated Reviews: Biochemistry
 الجزء والصفحة:
الجزء والصفحة:
 25-9-2021
25-9-2021
 6279
6279




+
-
20
Glycogenesis and Glycogenolysis Regulation
Because of the importance of maintaining blood glucose levels, the synthesis and degradation of its glycogen storage form are tightly regulated. In the liver, glycogenesis accelerates during periods when the body has been well fed, whereas glycogenolysis accelerates during periods of fasting. In skeletal muscle, glycogenolysis occurs during active exercise, and glycogenesis begins as soon as the muscle is again at rest. Regulation of synthesis and degradation is accomplished on two levels. First, glycogen synthase and glycogen phosphorylase are hormonally regulated (by covalent
phosphorylation/dephosphorylation) to meet the needs of the body as a whole. Second, these same enzymes are allosterically regulated (by effector molecules) to meet the needs of a particular tissue.
A. Covalent activation of glycogenolysis
The binding of hormones, such as glucagon or epinephrine, to plasma membrane G protein–coupled receptors ([GPCR] ) signals the need for glycogen to be degraded, either to elevate blood glucose levels or to provide energy for exercising muscle.
1. Protein kinase A activation: Binding of glucagon or epinephrine to their specific hepatocyte GPCR, or of epinephrine to a specific myocyte GPCR, results in the G protein–mediated activation of adenylyl cyclase. This enzyme catalyzes the synthesis of cyclic adenosine monophosphate (cAMP), which activates cAMP-dependent protein kinase A (PKA).
cAMP binds the two regulatory subunits of tetrameric PKA, releasing two individual catalytic subunits that are active (Fig. 1). PKA then phosphorylates several enzymes of glycogen metabolism, affecting their activity. [Note: When cAMP is removed, the inactive tetramer reforms.]
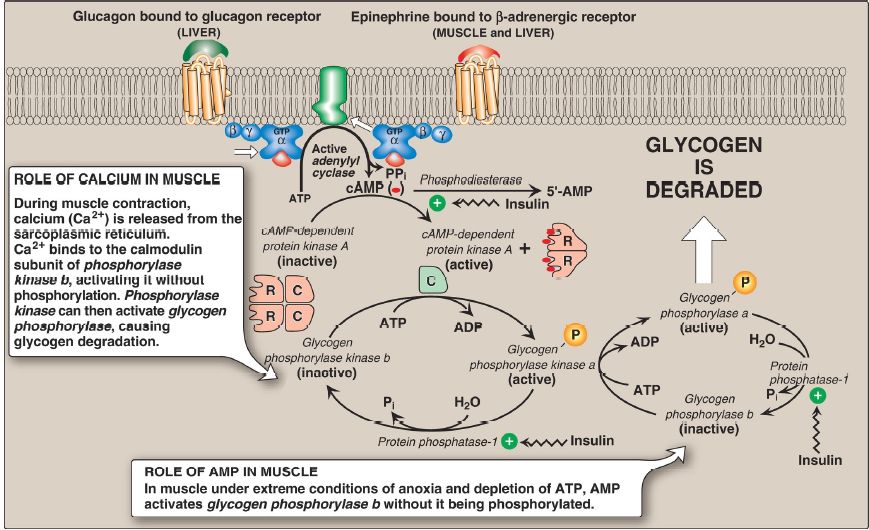
Figure 1: Stimulation and inhibition of glycogen degradation. AMP =adenosine monophosphate; cAMP = cyclic AMP; GTP = guanosine triphosphate; = phosphate; PPi = pyrophosphate; R = regulatory subunit; C = catalytic subunit.
2. Phosphorylase kinase activation: Phosphorylase kinase exists in two forms: an inactive “b” form and an active “a” form. Active PKA phosphorylates the inactive “b” form of phosphorylase kinase, producing the active “a” form (see Fig. 1).
3. Glycogen phosphorylase activation: Glycogen phosphorylase also exists in a dephosphorylated, inactive “b” form and a phosphorylated, active “a” form. Phosphorylase kinase a is the only enzyme that phosphorylates glycogen phosphorylase b to its active “a” form, which then begins glycogenolysis (see Fig. 1).
4. Signal amplification: The cascade of reactions described above activates glycogenolysis. The large number of sequential steps serves to amplify the effect of the hormonal signal (that is, a few hormone molecules binding to their GPCR result in a number of PKA molecules being activated that can each activate many phosphorylase kinase molecules).
This causes the production of many active glycogen phosphorylase a molecules that can degrade glycogen.
5. Phosphorylated state maintenance: The phosphate groups added to phosphorylase kinase and phosphorylase in response to cAMP are maintained because the enzyme that hydrolytically removes the phosphate, protein phosphatase-1 (PP1), is inactivated by inhibitor proteins that are also phosphorylated and activated in response to cAMP (see Fig. 1). [Note: PP1 is activated by a signal cascade initiated by insulin . Because insulin also activates the phosphodiesterase that degrades cAMP, it opposes the effects of glucagon and epinephrine.]
B. Covalent inhibition of glycogenesis
The regulated enzyme in glycogenesis, glycogen synthase, also exists in two forms, the active “a” form and the inactive “b” form. However, in contrast to phosphorylase kinase and phosphorylase, the active form of glycogen synthase is dephosphorylated, whereas the inactive form is phosphorylated at several sites on the enzyme, with the level of inactivation proportional to the degree of phosphorylation (Fig. 2). Phosphorylation is catalyzed by several different protein kinases in response to cAMP (for example, PKA and phosphorylase kinase) or other signaling mechanisms (see C. below). Glycogen synthase b can be reconverted to the “a” form by PP1. Figure 3 summarizes the covalent regulation of glycogen metabolism.

Figure 2: Hormonal regulation of glycogen synthesis. [Note: In contrast to glycogen phosphorylase, glycogen synthase is inactivated by phosphorylation.] cAMP = cyclic adenosine monophosphate; = phosphate; PPi = pyrophosphate; R = regulatory subunit; C = catalytic subunit; ADP = adenosine diphosphate.

Figure 3: Summary of the hormone-mediated covalent regulation of glycogen metabolism. cAMP = cyclic adenosine monophosphate; PKA = protein kinase A.
C. Allosteric regulation of glycogenesis and glycogenolysis
In addition to hormonal signals, glycogen synthase and glycogen phosphorylase respond to the levels of metabolites and energy needs of the cell. Glycogenesis is stimulated when glucose and energy levels are high, whereas glycogenolysis is increased when glucose and energy levels are low. This allosteric regulation allows a rapid response to the needs of a cell and can override the effects of hormone-mediated covalent regulation. [Note: The “a” and “b” forms of the allosteric enzymes of glycogen metabolism are each in an equilibrium between the R (relaxed, more active) and T (tense, less active) conformations . The binding of effectors shifts the equilibrium and alters enzymic activity without directly altering the covalent modification.]
1. Regulation in the well-fed state: In the well-fed state, glycogen synthase b in both liver and muscle is allosterically activated by glucose 6- phosphate, which is present in elevated concentrations (Fig. 4). In contrast, glycogen phosphorylase a is allosterically inhibited by glucose 6-phosphate, as well as by ATP, a high-energy signal. [Note: In liver, but not muscle, free glucose is also an allosteric inhibitor of glycogen phosphorylase a.]

Figure 4: Allosteric regulation of glycogenesis and glycogenolysis in liver (A) and muscle (B). P = phosphate; AMP = adenosine monophosphate.
2. Glycogenolysis activation by AMP: Muscle glycogen phosphorylase (myophosphorylase), but not the liver isozyme, is active in the presence of the high AMP concentrations that occur under extreme conditions of anoxia and ATP depletion. AMP binds to glycogen phosphorylase b, causing its activation without phosphorylation (see Fig. 1). [Note: Recall that AMP also activates phosphofructokinase-1 of glycolysis , allowing glucose from glycogenolysis to be oxidized.]
3. Glycogenolysis activation by calcium: Calcium (Ca2+) is released into the sarcoplasm in muscle cells (myocytes) in response to neural stimulation and in the liver in response to epinephrine binding to α1- adrenergic receptors. The Ca2+ binds to calmodulin (CaM), the most widely distributed member of a family of small, Ca2+-binding proteins. The binding of four molecules of Ca2+ to CaM triggers a conformational change such that the activated Ca2+–CaM complex binds to and activates protein molecules, often enzymes, that are inactive in the absence of this complex (Fig. 5). Thus, CaM functions as an essential subunit of many complex proteins. One such protein is the tetrameric phosphorylase kinase, whose “b” form is activated by the binding of Ca2+ to its δ subunit (CaM) without the need for the kinase to be phosphorylated by PKA. [Note: Epinephrine at β-adrenergic receptors signals through a rise in cAMP, not Ca2+ .]

Figure 5: Calmodulin mediates many effects of intracellular calcium (Ca2+). [Note: Ca2+ activates phosphorylase kinase in liver and muscles.]
a. Muscle phosphorylase kinase activation: During muscle contraction, there is a rapid and urgent need for ATP. It is supplied by the degradation of muscle glycogen to glucose 6-phosphate, which enters glycolysis. Nerve impulses cause membrane depolarization, which promotes Ca2+ release from the sarcoplasmic reticulum into the sarcoplasm of myocytes. The Ca2+ binds the CaM subunit, and the complex activates muscle phosphorylase kinase b (see Fig. 1).
b. Liver phosphorylase kinase activation: During physiologic stress, epinephrine is released from the adrenal medulla and signals the need for blood glucose. This glucose initially comes from hepatic glycogenolysis. Binding of epinephrine to hepatocyte α1-adrenergic GPCR activates a phospholipid-dependent cascade (see p. 205) that results in movement of Ca2+ from the ER into the cytoplasm. A Ca2+– CaM complex forms and activates hepatic phosphorylase kinase b. [Note: The released Ca2+ also helps to activate protein kinase C that can phosphorylate (therefore, inactivate) glycogen synthase a.]
 الاكثر قراءة في الكيمياء الحيوية
الاكثر قراءة في الكيمياء الحيوية
 اخر الاخبار
اخر الاخبار
اخبار العتبة العباسية المقدسة

الآخبار الصحية


مواضيع ذات صلة
 قسم الشؤون الفكرية يصدر كتاباً يوثق تاريخ السدانة في العتبة العباسية المقدسة
قسم الشؤون الفكرية يصدر كتاباً يوثق تاريخ السدانة في العتبة العباسية المقدسة "المهمة".. إصدار قصصي يوثّق القصص الفائزة في مسابقة فتوى الدفاع المقدسة للقصة القصيرة
"المهمة".. إصدار قصصي يوثّق القصص الفائزة في مسابقة فتوى الدفاع المقدسة للقصة القصيرة (نوافذ).. إصدار أدبي يوثق القصص الفائزة في مسابقة الإمام العسكري (عليه السلام)
(نوافذ).. إصدار أدبي يوثق القصص الفائزة في مسابقة الإمام العسكري (عليه السلام)



















