
Sigma Factors Can Be Organized into Cascades
 المؤلف:
JOCELYN E. KREBS, ELLIOTT S. GOLDSTEIN and STEPHEN T. KILPATRICK
المؤلف:
JOCELYN E. KREBS, ELLIOTT S. GOLDSTEIN and STEPHEN T. KILPATRICK
 المصدر:
LEWIN’S GENES XII
المصدر:
LEWIN’S GENES XII
 الجزء والصفحة:
الجزء والصفحة:
 6-5-2021
6-5-2021
 2203
2203
Sigma Factors Can Be Organized into Cascades
KEY CONCEPTS
- A cascade of sigma factors is created when one sigma factor is required to transcribe the gene encoding the next sigma factor.
- The early genes of phage SPO1 are transcribed by host RNA polymerase.
- One of the early genes encodes a sigma factor that causes RNA polymerase to transcribe the middle genes.
- Two of the middle genes encode subunits of a sigma factor that cause RNA polymerase to transcribe the late genes.
As in E. coli, sigma factors are used extensively to control initiation of transcription in the bacterium Bacillus subtilis. The B. subtilis genome encodes at least 18 different sigma factors, compared to the 7 found in E. coli. Larger numbers of sigma factors than in E. coli are not unusual. In fact, the Streptomyces coelicolor genome encodes more than 60!
In B. subtilis, some of the sigma factors are present in vegetative cells, whereas others are produced only in the special circumstances of phage infection or during the change from vegetative growth to sporulation. The major RNA polymerase engaged in normal vegetative growth contains the same subunits and has the same overall structure as that of E. coli, α2 ββ′ωσ, but in addition it has another subunit called δ. Its major sigma factor (σA ) recognizes promoters with the same consensus sequences used by the E. coli enzyme under direction from σ70 . Alternative RNA polymerases containing different sigma factors are found in much smaller amounts and recognize promoters with different consensus sequences in the −35 and −10 regions.
Transitions from expression of one set of genes to another set are a feature of bacteriophage infection. This is the case in B. subtilis infection by the phage SPO1, as it is in E. coli infection by phages such as T7, N4, or Φλ. In all but the very simplest cases, the development of the phage involves shifts in the pattern of transcription during the infective cycle. These shifts may be accomplished by the synthesis of a phage-encoded RNA polymerase or by the efforts of phage-encoded ancillary factors that control the bacterial RNA polymerase. During infection of B. subtilis by phage SPO1, the different stages of infection are controlled via the production of new sigma factors. The infective cycle of SPO1 has three stages of gene expression. Immediately on infection, the early genes of the phage are transcribed. After 4 to 5 minutes, the early genes cease transcription and the middle genes are transcribed. At 8 to 12 minutes, middle gene transcription is replaced by transcription of late genes.
The early genes are transcribed by the holoenzyme of the host bacterium. They are essentially indistinguishable from host genes whose promoters have the intrinsic ability to be recognized by the RNA polymerase α2ββ′ωσA .
Expression of phage genes is required for the transitions to middle and late gene transcription. Three regulatory genes—28,33, and 34 —control the course of transcription. Their functions are summarized in FIGURE 1. The pattern of regulation resembles a cascade, in which the host enzyme transcribes an early gene whose product is needed to transcribe the middle genes. After this transcription, two of the middle genes code for products that are needed to transcribe the late genes.
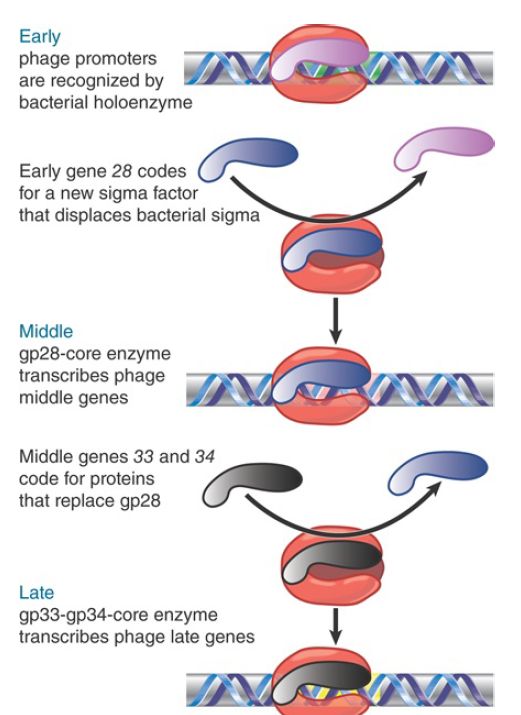
FIGURE 1.Transcription of phage SPO1 genes is controlled by two successive substitutions of the sigma factor that change the initiation specificity.
Mutants in the early gene 28 cannot transcribe the middle genes. The product of gene 28 (called gp28) is a 26-kD protein that replaces the host sigma factor on the core enzyme. This substitution is the sole event required to make the transition from early to middle gene expression. It creates a holoenzyme that can no longer transcribe the host genes but instead specifically transcribes the middle genes. It is not known how gp28 displaces σ43 or what happens to the host sigma polypeptide.
Two of the middle genes are involved in the next transition. Mutations in either gene 33 or 34 prevent transcription of the late genes. The products of these genes form a dimer that replaces gp28 on the core polymerase. Again, it is not known how gp33 and gp34 exclude gp28 (or any residual host σA ), but once they have bound to the core enzyme, they are able to initiate transcription only at the promoters for late genes.
The successive replacements of sigma factor have dual consequences. Each time the subunit is changed the RNA polymerase becomes able to recognize a new class of genes and it no longer recognizes the previous class. These switches therefore constitute global changes in the activity of RNA polymerase.
 الاكثر قراءة في مواضيع عامة في الاحياء الجزيئي
الاكثر قراءة في مواضيع عامة في الاحياء الجزيئي
 اخر الاخبار
اخر الاخبار
اخبار العتبة العباسية المقدسة




















